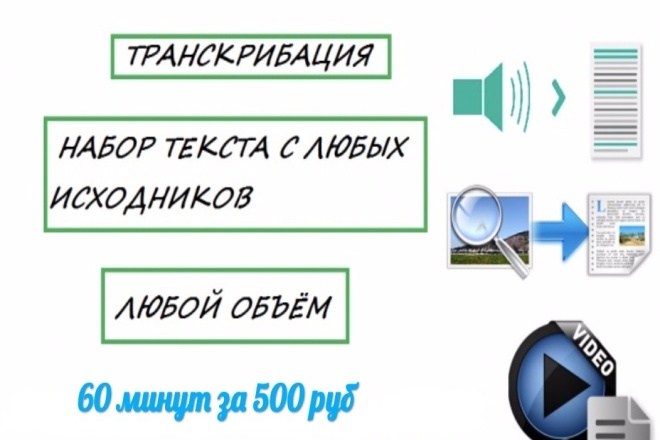
Биржа транскрибации: Заработок на транскрибации: лучшие сайты для транскрибаторов
Транскрибация за 1500 рублей | 19 объявлений в разделе расшифровка аудио
Сторимейкер
В команду SMM-агентства на постоянную работу требуется сторимейкер. В ваши обязанности будет входить: — Составление контент-плана — Создание сторис — Работа в системе trello (обучим) — Присутствие на общих…
Ирина Е.
Составить дизайн проект кухни
Проконсультировать по кухням, по материалам, нарисовать по моим размерам, сделать это так как делают в мебельных магазинах
Анастасия В.
Перенести иноформацию с сайта на стандатный шаблон…
Нужно перенести информацию и картинки с саита на шаблон , чтобы получился красивый магазин. Сайт одностраничный. Информации не много.
Галина М.
Архитектурный проект
Требуется проект каркасного дома 6х4 и беседки из бруса 6х4 с расчетом материалов.
Алексей
Задача, написать христианскую картину. Как будто под лучем света Иисуса стоят несколько человек. Спасая людей на земле. Картина должна иметь благодарственный характер . Предложения и суммы предлагайте
Дмитрий Р.
Страница не найдена — Интернет и деньги онлайн
Фриланс
Фрилансер (freelancer) это тоже, что и свободный художник или работник.
Подработка
На данный момент (2020 год) сайт Otzovik.com предоставляет самые лучшие условия и возможности для
Подработка
В начале трудового пути в интернете сложно оценить свои таланты и способности, особенно, если
Фотографом, как известно, может быть каждый. Это расхожее мнение сегодня, когда в интернете лидирует Instagram.
Фриланс
Биржа eTXT.ru — позиционирует себя как биржа для тех, кто умеет и любит писать
Фриланс
Вы уже давно задумываетесь о заработке в интернете? Возможно, вам не интересны маленькие суммы,
Как заработать на транскрибации видео или аудио: примеры, советы, личный опыт
- Светлана Билецкая
- 22.
 11.2020
11.2020 - Комментариев нет
Сегодня мы продолжаем тему “Дохода через интернет” и рассмотрим такой вариант заработка как “транскрибация” аудио и видео.
Для начала, давайте вспомним, что такое транскрибация.
Транскрибация — это перевод речи из аудио или видео в текст.
В вопросе заработка на транскрибации у меня есть две новости – хорошая и плохая.
Плохая состоит в том, что услуги по транскрибации стоят не дорого и, соответственно, доходы не большие.
Хорошая новость состоит в том, что этот вид дохода не требует особенных навыков.
Для того, чтобы заработать на транскрибации важны два ключевых момента:
- Знать, где искать заказчиков
- Уметь ее делать быстро
И в сегодняшнем посте я поделюсь такими вопросами:
- Как делать транскрибацию аудио и видео быстро: полезные советы, сайты, программы
- Где искать Заказчиков по транскрибации видео или аудио
- Примеры заказов по транскрибации
Как делать транскрибацию аудио и видео быстро: полезные советы, сайты, программы
Когда я только начинала искать и изучать разные варианты дохода через интернет, я пробовала многие задачи: транскрибацию, наполнение интернет-магазинов, сбор информации и в результате нашла себя в видеомонтаже. Сейчас у меня есть сайт по видеомонтажу, ютуб каналы и постоянные клиенты. Транскрибацией больше не занимаюсь.
Но когда-то этот опыт дал мне немного уверенности в том, что можно зарабатывать через интернет и клиентов по транскрибации я находила на ворзилле.
Единственное “но”, тогда я не знала о фишках, которые помогают сделать транскрибацию быстрее и, соответственно, заработать больше. Если бы знала, то смогла бы быстрее делать заказы и увеличить доход.![]()
И если вы как раз ищете информацию по поводу того, как быстрее сделать транскрибацию, делюсь с вами 2 способами.
Детально об этом я рассказала и показала в посте “Как быстро сделать транскрибацию видео и аудио: сравнение 2 способов”, там есть описание, скрины и видео.
Очень надеюсь, что эта информация поможет вам ускорить процесс транскрибации.
Где искать заказчиков по транскрибации видео или аудио
Я искала заказы на воркзилле и именно там находила клиентов по транскрибации.
Биржи фриланса – это самый простой и безопасный способ найти заказчиков по транскрибации. Вы видите конкретную стоимость, которую готов заплатить заказчик и объем работы, который нужно выполнить.
Если речь идет о воркзилле, то там заказчик назначает вас исполнителей и после этого вы выполняете задачу. Когда заказ сдан, производится оплата. На разных биржах фриланса, разные условия взаимодействия заказчика и исполнителя.
На нашем сайте мы сделали подборку 10 бирж фриланса и удаленной работы. Их можно посмотреть ЗДЕСЬ.
Важно! Я поделилась не просто списком сайтов, но и полезными советами по работе через биржи фриланса и удаленной работы
Примеры заказов по транскрибации
Сделала несколько скринов задач транскрибации на биржах фриланса.
На сегодня все!
Надеюсь, что информация для вас полезной.
Хороших вам доходов!
Рекомендую почитать:
Транскрибация и авторское право, DELETED — Advego.com
Тип текста: ЛюбойКопирайтингРерайтинг без источникаПеревод
Язык: ЛюбойRussian — РусскийEnglishGermany — DeutschSpanish — EspañolFrench — FrançaisChinese — 中国Ukrainian — УкраїнськаJapanese — 日本のPortuguese — PortuguêsPolish — PolskiItalian — ItalianoTurkish — TürkArabic — العربيةVietnamese — tiếng ViệtKorean — 한국의Urdu — اردوPersian — فارسیHindi — हिन्दीDutch — HollandskFinnish — suomalainenAnother language — другой язык
Категория: ЛюбаяБез категорииIT, софтАвиация, военная техника, ГОАвто, мотоАзартные игры, казино, покерБытовая техникаДизайн и предметы интерьераДомашние животныеДомашние растения, цветы, растительный мирЗакон и ПравоИгрушки, товары для детейИнтернет-маркетинг, SEO, SMM, создание сайтовИстория, религия, традиции и обрядыКиноКомпьютерные игры, видеоигры и приставкиКрасота и здоровье, питание, диеты, фитнесКулинарияКультура и искусствоЛандшафтный дизайн и архитектураМатериалы 18+Мебель и аксессуарыМедицина, лечение и профилактика болезнейМобильные игры и приложенияМода и СтильМузыкаНаука, открытия, высокие технологииНедвижимостьНепознанное: фэн-шуй, астрология, гороскопыОбразование, учеба, тренингиОтдых, активные игры, охота и рыбалкаОтношения, знакомства, личная жизньПолиграфия, рекламная продукция, маркетингПолитика: аналитика и обзорыПраздники и торжества, свадьбаПрирода и экологияПромышленность и оборудованиеПсихологияРабота и карьера, фрилансРемонт и обустройствоРукоделие, хобби, handmadeСад и огород, сельское хозяйствоСемья, воспитание детей, беременность и родыСобственный бизнес, ForexСпорт и спортивный инвентарь, велотехникаСтихи и поздравленияСтроительный инструмент и материалы, садовая техникаСтроительство домов, дачное хозяйствоТуризм, достопримечательностиУслуги и сервисФинансы, банки и кредиты, экономикаФототехника, искусство фотографииЭлектроника: гаджеты, мобильные телефоны, компьютеры, телевизорыЮмор
Как упростить процесс транскрибации (расшифровки аудио- и видеозаписей)
Транскрибация — это расшифровка аудио— и видеоматериалов, их содержимое, переведённое в текст. То, о чём говорится в таких материалах, дословно или с определённой степенью цензуры отображается в текстовом документе. В этой статье будем говорить о способах упрощения транскрибации на компьютере. Казалось бы, что сложного в процессе расшифровки? Берёшь удобный медиаплеер, периодически приостанавливаешь запись и переписываешь услышанное.
То, о чём говорится в таких материалах, дословно или с определённой степенью цензуры отображается в текстовом документе. В этой статье будем говорить о способах упрощения транскрибации на компьютере. Казалось бы, что сложного в процессе расшифровки? Берёшь удобный медиаплеер, периодически приостанавливаешь запись и переписываешь услышанное.
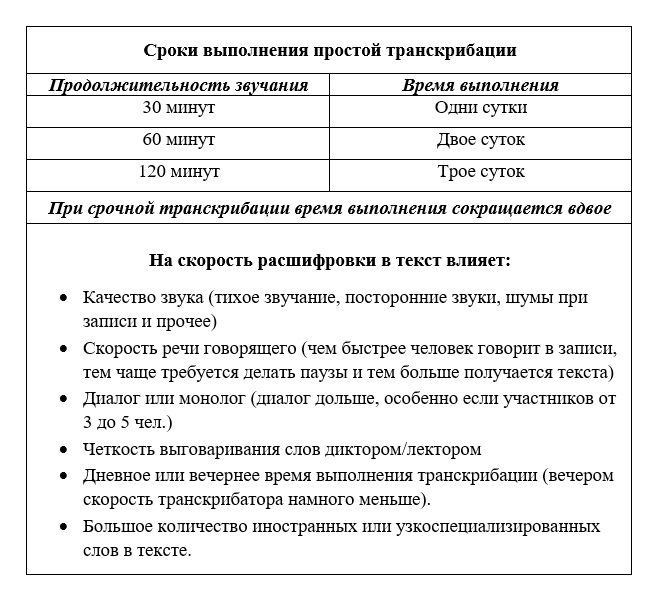
1. Сложности транскрибации
Но нет, всё не так просто. Иначе заинтересованные в такой услуге лица не искали бы удалённых сотрудников на биржах фриланса.
Во-первых, не каждый сможет выполнить такую работу. Нужен выдержанный, усидчивый, способный к длительной концентрации внимания человек.
Во-вторых, не всегда для расшифровки поставляется качественный исходник, а это, соответственно, усложняет процесс транскрибации. В таких случаях исполнитель должен быть ещё и технически подкован, чтобы решить вопрос с устранением помех записи.
В-третьих, если речь идёт о создании расшифровки для публичных целей, за выполнение работы должен взяться человек грамотный, со знанием речи. В качестве исходника редко когда поставляются студийные записи с чёткой речью диктора. Чаще расшифровывать приходится рабочие материалы — интервью, семинары, вебинары, лекции, диктовки, прослушки, протоколы заседаний и т.п. Вживую же люди говорят так, как они говорят. Исполнителю нужно ещё и перефразировать их речь – убрать диалект, междометия, слова-паразиты, мат, построить предложения грамотно, оставив суть сказанного той же.
Транскрибация
– процесс ресурсоёмкий. Постоянное переключение внимания от одной задачи к другой, от одного программного окна к другому быстро истощает силы. Можно ли как-то упростить этот процесс? Увы, идеального способа автоматизации этого процесса пока что не существует. Справиться с такой задачей может только человек. Но его работу можно упростить с помощью отдельных программных средств.2.
Для удобства работы с двумя программными окнами – медиаплеера и текстового редактора – в среде современных версий Windows можно прибегнуть к функции эффективного задействования пространства экрана Snap. Окно плеера крепим к одной части экрана, окно редактора – к другой.
Совет: при расшифровке аудиоматериалов лучше использовать плееры с крупными элементами управления, как, например, приложение «Музыка Groove», поставляемое вместе с Windows 10.
3. Штатный видеоплеер Windows 10
При транскрибации видеозаписей можно задействовать компактный режим штатного видеоплеера Windows 10 – приложения
Совет: если исходником является видео, чтобы не отвлекаться на видеоряд, можно извлечь звук в аудиофайл. И работать уже с ним. Такие операции умеют проводить программы-медиаконвертеры, к примеру, бесплатные Convertilla и Freemake Video Converter. А программы типа Adobe Audition или её бесплатного аналога Audacity помогут улучшить качество звучания, если исходное оставляет желать лучшего.
4. Сторонние видеоплееры
Сторонние видеоплееры для Windows, такие как, например, KMPlayer, GOM, VLC, предложат больший функционал, который, в частности, может быть задействован при транскрибации:
• Настраиваемые шаги вперёд и назад;
• Регулировка скорости воспроизведения;
• Установка закладок;
• Настраиваемая панель элементов управления;
• Настраиваемые горячие клавиши.
Совет: некоторые исполнители предпочитают не приостанавливать исходник, а замедлять его воспроизведение для написания под диктовку. Это не лучшая идея в целях оптимизации транскрибации. При написании диктанта важны паузы, разграничивающие даже части предложений. Плюс к этому, наше сознание входит в резонанс с замедленным воспроизведением, а это, соответственно, тормозит работу ума и набор текста.
Это не лучшая идея в целях оптимизации транскрибации. При написании диктанта важны паузы, разграничивающие даже части предложений. Плюс к этому, наше сознание входит в резонанс с замедленным воспроизведением, а это, соответственно, тормозит работу ума и набор текста.
5. Программа Express Scribe
Express Scribe – программа, специально созданная для транскрибации. В верхней части её интерфейса находится поле для добавления медиафайлов, в нижней – проигрыватель и текстовый редактор. В числе её преимуществ:
• Единое окно для работы с исходниками и текстом;
• Поддержка большого перечня медиаформатов;
• Функции замедления и ускорения проигрываемых файлов;
• Перемотка;
• Улучшение качества звучания;
• Горячие клавиши, а также возможность задействования для отдельных функций ножных педалей геймерского руля;
• Сохранение исходников и их расшифровки как проектов программы;
• Экспорт набранного текста в Microsoft Word.
Недостатки Express Scribe – отсутствие поддержки русского языка интерфейса и активное навязывание создателями программы платных её редакций при работе с бесплатной.
6. Сервисы голосового ввода данных
Веб-сервисы, предлагающие возможность голосового ввода данных – пожалуй, самый оптимальный способ упрощения транскрибации. Особенно подойдёт тем, чья скорость печати оставляет желать лучшего. Принцип работы здесь прост: запоминаем отрывки текста, повторяем их в микрофон и попутно проводим коррекцию текста – исправляем ошибки и вставляем знаки препирания.
Механизмом распознавания речи оснащён веб-сервис «Google Документы». Создаваемые в этом онлайн-редакторе документы можно надиктовывать с использованием инструмента «Голосовой ввод».
Инструмент поддерживает огромный перечень языков, включая русский. Однако он не предусматривает настройку автоотключения ввода данных. Если не говорить в микрофон пару секунд, голосовой ввод отключается. И его нужно постоянно включать вручную.
И его нужно постоянно включать вручную.
Другой веб-сервис – «Голосовой блокнот Speechpad.ru» — будет в разы эффективнее, поскольку это узкопрофильный проект, и распознавание речи является его основным направлением. Для ввода голосовых данных в форме сервиса необходимо нажать кнопку включения записи. Текст правится в результирующем поле. Внизу этого поля присутствует кнопка «+Транскрибация».
Она добавляет в форму сервиса возможность воспроизведения аудио- и видеоисходников для работы в одном программном окне. Например, чтобы сделать расшифровку ролика на YouTube, в форме сервиса необходимо выбрать его медиатип и вставить в соответствующее поле ID (часть веб-адреса после знака «=»).
Сервис кроме русского поддерживает ещё несколько других языков.
Совет: Speechpad.ru умеет распознавать два знака препинания – точку и запятую. Их можно проговаривать.
Недостаток такого способа транскрибации – несовершенство механизма распознавания человеческой речи. Сколькие из мобильных пользователей, вдоволь наигравшись с «Ок, Google» при вводе поисковых запросов, пришли к тому, что проще отыскать что-то по старинке, с помощью клавиатуры. Какой бы сервис ни использовался, всё равно в процессе работы придётся делать паузы и отслеживать распознанный текст мелкими блоками.
7. Автоматизация транскрибации
Процесс голосового ввода данных при транскрибации можно упростить по максимуму, настроив системное звучание в качестве источника звукозаписи и, соответственно, убрав из цепочки действий проговаривание услышанного в микрофон. Естественно, в случае с проблемными исходниками такой вариант не подойдёт. В итоге получим нечитаемую белиберду, которую проще будет переписать заново, чем редактировать. Если же запись более-менее качественная, попробовать стоит.
Вызываем контекстное меню на значке звука в области задач Windows, выбираем «Звуки». В появившемся окошке с помощью опций в контекстном меню отключаем микрофон и включаем стереомикшер.
В появившемся окошке с помощью опций в контекстном меню отключаем микрофон и включаем стереомикшер.
Далее воспроизводим видео или аудио и расшифровываем его с помощью веб-сервиса Speechpad.ru.
Однако процесс всё равно нужно контролировать: вникать в суть информации, периодически приостанавливать воспроизведение для коррекции расшифровки, проставлять знаки препирания.
8. И самое главное …
Осуществлять транскрибацию на ноутбуке с его хрипящим динамиком проблематично. Однако в особо сложных случаях не поможет и хорошая аудиосистема. Для постоянной работы по расшифровке аудио лучше обзавестись наушниками. А если работа проводится путём распознавания речи, возможно, стоит присмотреть себе качественную гарнитуру Hands Free.
Расшифровка аудио: определение, особенности и программы
Расшифровка аудио – это процесс перевода речи из аудиоформата в текстовый. Иными словами его называют транскрибацией.
Расшифровке подлежат:
- Записи интервью и подкастов
- Аудиодорожки из видеороликов и телепередач
- Записи разговоров в Zoom и Skype
- Лекции, семинары, конференции
Существуют два способа расшифровки аудио в текст: ручная транскрибация специалистами-расшифровщиками и компьютерные программы и онлайн сервисы расшифровки. Рассмотрим особенности каждого из них подробнее.
Транскрибация специалистом
Расшифровщики способны печатать текст очень быстро, они внимательны к деталям. Перед расшифровкой специалист прослушивает запись целиком, чтобы определить ее тему, действующих лиц и канву разговора. Получив общее представление о записи, он приступает к ее переводу в текстовый формат.
Главный плюс расшифровки аудиозаписи и видеозаписи человеком – это то, что качество расшифровки будет максимально высоким, так как расшифровщик не только транскрибирует аудио в текст, но и может выступать в роли редактора.
Кроме того, программы расшифровки не смогут разделить текст на абзацы, и большинство из них не умеют ставить знаки препинания.
Минус ручной расшифровки в том, что она занимает много времени. Например, на расшифровку двухчасовой аудиозаписи у специалиста может уйти целый день. Однако качество финального текста будет на высоте.
Найти специалиста по транскрибации текста вы можете в нашем бюро переводов “ЛингваКонтакт”, а также на биржах фриланса, например на Workzilla, Kwork, text.ru. Мы советуем обращаться расшифровкой аудио к людям, если речь на записи не самого хорошего качества.
Программы для расшифровки аудио в текст
Существует немало онлайн программ для расшифровки аудио в текст. Если раньше качество их расшифровки оставляло желать лучшего, то с каждым годом процент ошибок, который они допускают, становится все меньше.
И все же любая программа для расшифровки аудио в текст, особенно бесплатная, не способна справиться с некоторыми аудиозаписями. Допустим, нужно переложить на текст оживленную дискуссию.
На записи участники перекрикивают друг друга, сбиваются с мысли. Восприятию звука могут помешать и посторонние шумы. Программа расшифровки аудиозаписи в текст не справится с такой сложной задачей: скорее всего придется редактировать запись вручную.
Базовые сервисы по расшифровке аудио: textforspeech.com, Блокнот для речевого ввода, dicatation.io, расширение “голосовой ввод текста” Google Chrome.
Продвинутые сервисы по транскрибации с бесплатным пробным периодом: trint.com, sonix.ai, otter.ai, happyscribe.com.
Вам решать, какой способ расшифровки использовать. Если решите нанять специалиста по расшифровке текста, советуем обратиться к нашим транскрибаторам. Наши специалисты проставят таймкоды, отформатируют и отредактируют текст. На руки Вы получите готовый к использованию материал.
Желаем хорошего дня!
Заказы на транскрибацию — Удаленная работа в интернете
Доброго здоровья, уважаемый читатель журнала «Web4job.![]() ru”! В этой статье мы поговорим о том, как получить заказы на транскрибацию, кто пользуется этой услугой и какие навыки необходимы при этой работе.
ru”! В этой статье мы поговорим о том, как получить заказы на транскрибацию, кто пользуется этой услугой и какие навыки необходимы при этой работе.
Заказы на траскрибацию
Содержание статьи:
Как мы уже знаем, что транскрибация – это перевод аудио и видео материалов в текстовый формат.
Это один из видов удаленной работы. В штате компаний обычно такой должности нет.
Постоянные клиенты, заказывающие транскрибацию, это:
Инфобизнесмены. Сейчас проводится много вебинаров, тренингов, конференций, темы которых записываются на аудио или видео формат.
В последующем они переводятся в текст, и печатаются книги, которые продаются, как новый инфопродукт.
Многие крупные компании проводят для рекламных агентств конференции, позже информация переводится в текст и публикуется на корпоративных сайтах.
В судах также ведутся аудиозаписи, которые в последующем переводятся в текст и сдаются в архив для хранения.
Радио и телевидение регулярно нуждаются в новостной информации.
Журналисты перерабатывают информацию в интересную статью.
Владельцы сайтов берут интервью и размещают его как в виде аудио и видеоматериала на сайте, так и в текстовом формате.
Писатели пишут свои мысли, а некоторые и книги, проговаривая их на диктофон, затем переводят их в текст.
Требования к работе
Чтобы делать транскрибацию, человек должен знать, какие требования к этой работе предъявляются и какими знаниями нужно обладать:
Грамотность. При переводе текста необходимо знать орфографию, стилистику, пунктуацию.
При редактировании нужно будет убирать слова-паразиты, править ошибки.
У исполнителя должен быть хороший слух и память. Если при прослушивании плохо запоминается тест, очень сложно будет работать:
придется часто останавливать запись, перематывать ее назад. Это неудобно при работе с большим объемом информации.
Необходимо быть усидчивым и терпеливым. Некоторые записи продолжаются по несколько часов, и, кроме того, что их нужно перевести,
необходимо грамотно оформить текст и устранить все ошибки.
Иметь навык быстрой печати, желательно уметь печатать слепым методом. Это также большое преимущество.
Чем быстрее печатаете, тем больше информации обрабатываете и больше получаете за свою работу.
Крепкие нервы. Они понадобятся при транскрибации записей из судов, где разбираются разные, в том числе, тяжелые жизненные ситуации – изнасилования, убийства.
Чтобы понять, подходит вам этот вид заработка или нет, можно просто взять любой видеоматериал и попробовать перевести его в текст.
Если вам это понравится, тогда можно начать брать заказы на биржах и пробовать на них зарабатывать.
Посмотрим видео о том, как получить заказы на транскрибацию
Заработок на транскрибации
Заработок на транскрибации. Транскрибация аудио в текст
Заключение
В этой статье мы поговорили о том, как получить заказы на транскрибацию и кто пользуется этой услугой.
Надеюсь, статья оказалась полезной. Если возникли вопросы, можете задать их через форму комментариев под этой статьей.
Также буду признательна, если поделитесь статьей со своими друзьями в социальных сетях.
Понравилось? Поделитесь с друзьями!
Получите высокооплачиваемую интернет-профессию!
определение, произношение, транскрипция, словоформы, примеры
существительное
— химический процесс, в котором один атом, ион или группа меняются местами с другим— взаимное выражение взглядов (особенно неприятных)
между ними произошел ожесточенный обмен
Адаму было обещано бессмертие в обмен на его непослушание
допустимых вычетов убытков от продажи или обмена собственности
 : центральный)
: центральный) — рабочее место для купли-продажи; открыт только для участников
— (спорт) непрерывная последовательность нескольких последовательных ударов (син.: ралли)
— взаимный перевод эквивалентных денежных сумм (особенно валют разных стран) (син .: обмен)
— акт замены одной вещи или человека на место другого: (синоним: замена, замена)
— (шахматы) получение (или потеря) ладьи взамен коня или слона
черные проиграли размен
эндшпиль начался после размена ферзей
глагол
— отдавать и получать друг от друга (син.— обменять или заменить другим, обычно того же типа или категории (синхр .: изменить, перебросить, преобразовать)
— переключить, поменять на новый порядок или последовательность (синхр .: переключить, переключить )
— сдать одного и получить другого, примерно эквивалентно
заключенных по обмену
сотрудника по обмену между филиалами компании
— обменять штраф на менее суровый (синоним: перебросить, преобразовать)
Дополнительные примеры
Мы обменялись адресами и пообещали, что будем часто писать друг другу.
Эти купоны можно обменять на еду.
Я хочу обменять этот свитер на меньший.
Между двумя лидерами состоялся краткий обмен мнениями.
Я хочу обменять это платье на платье большего размера.
Обменялись ударами.
Я предложил покрасить кухню в обмен на недельное проживание.
Четыре мои кассеты для вашего компакт-диска Мадонны — это честный обмен.
Диджея уволили после жаркой беседы (= очень злой беседы) в прямом эфире с приглашенным слушателем.
Политика магазина не допускает возврата или обмена.
В большинстве столиц есть обширные возможности обмена.
Я здесь на один срок, по обмену с доктором.Фишер.
Кейт в Нью-Йорке по обмену сотрудниками, чтобы пройти дополнительное обучение.
Обмениваемся подарками на Рождество.
В конце игры игроки традиционно обмениваются футболками друг с другом.
Формы слова
глагол
I / you / we / they: exchange
he / she / it: обмен
причастие настоящего времени: обмен
прошедшее время: обмен
причастие прошедшего времени: обмен
существительное
единственное число: обмен
множественное число: обмен
Разрешить пользователям видеть стенограмму голосовой почты в Exchange Online
- 7 минут на чтение
В этой статье
Предварительный просмотр голосовой почты — это функция, доступная пользователям, которые получают сообщения голосовой почты из единой системы обмена сообщениями (UM).Предварительный просмотр голосовой почты расширяет возможности существующей голосовой почты единой системы обмена сообщениями, предоставляя текстовую версию аудиозаписей. Текст голосовой почты отображается в сообщениях электронной почты в Microsoft Outlook 2010 или более поздней версии, Outlook в Интернете и в других поддерживаемых почтовых программах.
Нужно ли пользователям использовать определенную почтовую программу?
Нет. Предварительный просмотр голосовой почты включен в текст сообщения любой почтовой программы, включая мобильные программы. Хотя пользователи могут использовать другие почтовые программы для получения голосовых сообщений, Outlook и Outlook в Интернете обеспечивают более удобную работу.Например, в Outlook 2010 и более поздних версиях при щелчке определенного слова в тексте предварительного просмотра голосовой почты начинается воспроизведение звука голосового сообщения с этого слова. Это полезно для прослушивания определенной части голосового сообщения.
Могут ли пользователи искать определенные сообщения голосовой почты?
Да. Слова и фразы в тексте предварительного просмотра голосовой почты автоматически индексируются, поэтому голосовые сообщения будут отображаться в результатах поиска. В Outlook 2010 и более поздних версиях или в Outlook в Интернете пользователи также могут использовать поле Audio Notes для добавления текста о голосовом сообщении.Эти заметки также включаются в поиск, чтобы упростить поиск сообщения.
Почему эта функция называется «Предварительный просмотр голосовой почты»?
Важно правильно формировать ожидания пользователей. Предварительный просмотр голосовой почты не обязательно создает текст, который совпадает с тем, что звонящие говорят в своих голосовых сообщениях. На самом деле, это обычно в чем-то неточно. Назвать это транскрипцией означало бы получить более совершенный результат, чем обычно достижимый. Предварительный просмотр предполагает, что читатель должен понимать суть голосового контента, что ближе к реальным возможностям функции.
Что делает текст предварительного просмотра голосовой почты более или менее точным?
Точность текста предварительного просмотра голосовой почты зависит от многих факторов, и иногда эти факторы невозможно контролировать. Однако текст предварительного просмотра голосовой почты будет более точным, если:
Вызывающий абонент оставляет простое голосовое сообщение, не содержащее сленговых терминов, технического жаргона или необычных слов или фраз.

Вызывающий абонент использует язык, который легко распознается и переводится системой голосовой почты.Как правило, голосовые сообщения, оставленные абонентами, которые не говорят слишком быстро или тихо и у которых нет сильного акцента, дают более точные предложения и фразы.
Голосовое сообщение не содержит фонового шума, эха, а звук не пропадает.
Какие языки можно использовать с предварительным просмотром голосовой почты?
Текст для предварительного просмотра голосовой почты доступен на следующих языках:
Если у вас локальное или гибридное развертывание единой системы обмена сообщениями, вы можете загрузить языковые пакеты единой системы обмена сообщениями из Центра загрузки Майкрософт.
Если у вас локальное или гибридное развертывание, после установки языкового пакета единой системы обмена сообщениями абонентские группы и автосекретаря можно настроить для использования выбранного вами языка. Для онлайн-клиентов вам не нужно устанавливать какие-либо языковые пакеты единой системы обмена сообщениями. Многие компании имеют только одну абонентскую группу единой системы обмена сообщениями. Единая система обмена сообщениями попытается создать предварительный просмотр голосовой почты на языке абонентской группы по умолчанию, но будет успешна только в том случае, если язык по умолчанию поддерживает предварительный просмотр голосовой почты. Абонентскую группу единой системы обмена сообщениями можно настроить для создания предварительных просмотров голосовой почты только на одном языке.
Чтобы настроить единую систему обмена сообщениями для предварительного просмотра голосовой почты на языке, отличном от американского, выполните следующие действия:
Убедитесь, что предварительный просмотр голосовой почты поддерживается на языке, который вы хотите использовать.
Если у вас локальное или гибридное развертывание, загрузите и установите соответствующий языковой пакет единой системы обмена сообщениями.
 При загрузке и установке языкового пакета не настраивается язык абонентской группы по умолчанию.
При загрузке и установке языкового пакета не настраивается язык абонентской группы по умолчанию.Настройте абонентскую группу с языком, который будет использоваться для предварительного просмотра голосовой почты.Дополнительные сведения см. В разделе Установка языка по умолчанию для абонентской группы.
Как в предварительном просмотре голосовой почты отображается текст на поддерживаемых языках, зависит от типа отправленного голосового сообщения. Есть два типа:
Голосовые сообщения, которые записываются, когда пользователь не отвечает на свой телефон
Для этих сообщений язык, используемый для предварительного просмотра голосовой почты, определяется разговорным языком вызывающего абонента и тем, поддерживается ли этот язык.Например, если вызывающий абонент оставляет голосовое сообщение на итальянском языке, текст предварительного просмотра голосовой почты будет отображаться на итальянском языке, если итальянский язык был настроен в абонентской группе. Однако, если вызывающий абонент оставляет сообщение на японском языке, текст предварительного просмотра голосовой почты не будет включен в сообщение, поскольку японский язык недоступен.
Голосовые сообщения, отправленные пользователем голосового доступа к Outlook
Для сообщений, отправленных пользователем голосового доступа к Outlook, язык, используемый для предварительного просмотра голосовой почты, контролируется администратором голосовой почты.Таким образом, текст предварительного просмотра голосовой почты будет на том же языке, что и система голосовой почты. Однако, если вызывающий абонент, говорящий на языке, который не поддерживается для предварительного просмотра голосовой почты, использует голосовой доступ к Outlook, чтобы оставить сообщение, текст предварительного просмотра голосовой почты не будет включен в сообщение. Дополнительные сведения о голосовом доступе к Outlook см.
 В разделе Настройка голосового доступа к Outlook.
В разделе Настройка голосового доступа к Outlook.
Знает ли единая система обмена сообщениями, когда предварительный просмотр голосовой почты неточен?
Уровень достоверности определяется для каждого предварительного просмотра голосовой почты, включенного в голосовое сообщение.Система голосовой почты измеряет, насколько хорошо звуки в записи соответствуют словам, числам и фразам. Если совпадения обнаруживаются легко, уровень достоверности высокий. Более высокий уровень уверенности обычно связан с более высокой точностью.
Если уровень достоверности определен ниже определенного значения, фраза Предварительный просмотр голосовой почты (достоверность низкая) включается над текстом предварительного просмотра голосовой почты. Если уровень достоверности низкий, вероятно, текст предварительного просмотра голосовой почты будет неточным.
Единая система обмена сообщениямииспользует автоматическое распознавание речи (ASR) для расчета достоверности при предварительном просмотре, но у нее нет способа определить, какие слова неправильные, а какие правильные.
Однако единая система обмена сообщениями пытается научиться повышать точность предварительного просмотра голосовой почты. Например, он пытается сопоставить номер телефона вызывающего абонента (если он указан) с личными контактами пользователя и адресной книгой вашей организации или контактами из социальных сетей. Если единая система обмена сообщениями находит совпадение, оно будет включать имя вызывающего абонента вместе со стандартными списками имен и слов при запуске ASR для записи голоса.
Можно ли использовать предварительный просмотр голосовой почты, если он не совсем точен?
Пользователи могут лучше понять предварительный просмотр голосовой почты, если они не будут пытаться читать предварительный просмотр слишком внимательно, слово за словом. Вместо этого им следует искать имена, номера телефонов и фразы, такие как «Перезвоните мне» или «Мне нужно поговорить», которые могут помочь понять цель звонка.
не будет точно диктовать сообщения, но он может помочь пользователям ответить на такие вопросы, как следующие:
Связано ли это голосовое сообщение с моей работой?
Это голосовое сообщение важно для меня?
Позвонивший оставил номер? Отличается ли он от любых цифр, которые я мог для них перечислить?
Считает ли вызывающий абонент это голосовое сообщение срочным?
Следует ли мне выйти из встречи, чтобы перезвонить этому человеку?
Я ожидал звонка, чтобы подтвердить мой запрос.Это звонок для подтверждения?
Можно ли включить или выключить предварительный просмотр голосовой почты?
Да. Если вы включили предварительный просмотр голосовой почты, пользователи могут включить или выключить его с помощью Outlook 2010 или более поздней версии или Outlook в Интернете. Однако язык абонентской группы должен поддерживать предварительный просмотр голосовой почты, и должен быть установлен языковой пакет единой системы обмена сообщениями для этого языка.
Хотя настройки предварительного просмотра голосовой почты одинаковы, независимо от того, использует ли пользователь Outlook 2010 или более позднюю версию или Outlook в Интернете, они будут обращаться к ним по-разному:
Outlook в Интернете
Чтобы получить доступ к параметрам предварительного просмотра голосовой почты в Outlook в Интернете, пользователи щелкните Параметры > телефон > Голосовая почта .На странице Голосовая почта настройки доступны в разделе Предварительный просмотр голосовой почты .
По умолчанию, оба параметра предварительного просмотра голосовой почты доступны, если пользователю включена единая система обмена сообщениями. Если абонентская группа единой системы обмена сообщениями настроена на использование языкового пакета единой системы обмена сообщениями, который поддерживает предварительный просмотр голосовой почты, единая система обмена сообщениями будет создавать предварительные просмотры голосовой почты для пользователей, когда:
Вызывающий абонент оставляет сообщение голосовой почты, потому что пользователь не отвечает на его телефон.

Пользователь с включенной поддержкой единой системы обмена сообщениями входит в систему голосового доступа к Outlook и записывает голосовое сообщение для одного или нескольких получателей.
Когда вызывающий абонент оставляет голосовое сообщение и выбирается Включить текст предварительного просмотра в голосовые сообщения, которые я получаю , единая система обмена сообщениями создает предварительный просмотр голосовой почты в сообщении электронной почты, прикрепляет аудиофайл и отправляет его в почтовый ящик получателя. Вы можете отключить этот параметр, если язык, настроенный в абонентской группе, не включает поддержку предварительного просмотра голосовой почты, и вы не хотите, чтобы предварительный просмотр голосовой почты включался в сообщения голосовой почты.
Когда пользователи входят в голосовой доступ к Outlook и отправляют голосовое сообщение другому пользователю, они могут захотеть снять флажок Включить текст предварительного просмотра в голосовые сообщения, которые я отправляю через голосовой доступ к Outlook .Например, они могут захотеть сделать это, если они отправляют голосовые сообщения на языке, который не поддерживает предварительный просмотр голосовой почты, или если они не хотят включать предварительный просмотр голосовой почты в голосовое сообщение, потому что оно слишком длинное.
Обмен гистонов, структура хроматина и регуляция транскрипции
Йонкерс, И. и Лис, Дж. Т. Освоение элонгации транскрипции с помощью РНК-полимеразы II. Nature Rev. Mol. Cell Biol. http://dx.doi.org/10.1038/nrm3953 (2015).
Sainsbury, S., Bernecky, C. & Cramer, P. Структурные основы инициации транскрипции с помощью РНК-полимеразы II. Nature Rev. Mol. Cell Biol. http://dx.doi.org/10.1038/nrm3952 (2015).
Пурра, О. и Доменико Либри, Д. Терминация транскрипции и контроль транскриптома: почему, где и как остановиться. Nature Rev. Mol. Cell Biol. http://dx.doi.org/10.1038/nrm3943 (2015).
Nature Rev. Mol. Cell Biol. http://dx.doi.org/10.1038/nrm3943 (2015).
Люгер, К., Дечасса, М. Л. и Треметик, Д. Дж. Новое понимание структуры нуклеосом и хроматина: упорядоченное состояние или неупорядоченное дело? Nature Rev. Mol. Cell Biol. 13 , 436–447 (2012).
Артикул CAS Google Scholar
Тальберт, П. Б. и Хеникофф, С. Варианты гистонов — древние художники обертывания эпигенома. Nature Rev. Mol. Cell Biol. 11 , 264–275 (2010).
Артикул CAS Google Scholar
Корнберг, Р. Д. Структура хроматина: повторяющаяся единица гистонов и ДНК. Наука 184 , 868–871 (1974).
Артикул CAS PubMed Google Scholar
Marzluff, W.F., Gongidi, P., Woods, K.R., Jin, J. & Maltais, L.J. Гены гистонов, зависимые от репликации человека и мыши. Genomics 80 , 487–498 (2002).
Артикул CAS PubMed Google Scholar
Ahmad, K. & Henikoff, S. Вариант гистона h4.3 маркирует активный хроматин путем независимой от репликации сборки нуклеосом. Мол. Ячейка 9 , 1191–1200 (2002).
Артикул CAS PubMed Google Scholar
Вебер, К.М. и Хеникофф, С. Варианты гистонов: динамическая пунктуация в транскрипции. Genes Dev. 28 , 672–682 (2014).
Артикул CAS PubMed PubMed Central Google Scholar
Tagami, H., Ray-Gallet, D., Almouzni, G. & Nakatani, Y. Комплексы гистонов h4.1 и h4.3 опосредуют пути сборки нуклеосом, зависимые или независимые от синтеза ДНК. Cell 116 , 51–61 (2004). Очистка человеческих вариантных гистонов, проведенная в этом исследовании, помогла определить новые профили взаимодействия, особенно в отношении гистоновых шаперонов.![]()
Артикул CAS PubMed Google Scholar
Stoler, S., Keith, KC, Curnick, KE & Fitzgerald-Hayes, M. Мутация в CSE4 , важном гене, кодирующем новый связанный с хроматином белок у дрожжей, вызывает нерасхождение хромосом и клеточный цикл остановка при митозе. Genes Dev. 9 , 573–586 (1995).
Артикул CAS PubMed Google Scholar
Schenk, R., Jenke, A., Zilbauer, M., Wirth, S. & Postberg, J. h4.5 представляет собой новый гоминид-специфичный вариант гистона h4, который специфически экспрессируется в семенных канальцах яички человека. Хромосома 120 , 275–285 (2011).
Артикул CAS PubMed Google Scholar
Монтелье, Э.и другие. Переход хроматина к нуклеопротамину контролируется вариантом Th3B гистона h3B. Genes Dev. 27 , 1680–1692 (2013).
Артикул CAS PubMed PubMed Central Google Scholar
Люгер, К., Мадер, А. В., Ричмонд, Р. К., Сарджент, Д. Ф. и Ричмонд, Т. Дж. Кристаллическая структура ядерной частицы нуклеосомы с разрешением 2,8 Å. Nature 389 , 251–260 (1997). Это первое сообщение о кристаллической структуре нуклеосом, которое подчеркивает важность модульной организации.
Артикул CAS PubMed Google Scholar
Smith, S. & Stillman, B. Поэтапная сборка хроматина во время репликации ДНК in vitro . EMBO J. 10 , 971–980 (1991).
Артикул CAS PubMed PubMed Central Google Scholar
Кулаева, О.I., Hsieh, F.K. и Studitsky, V.M. Комплексы РНК-полимеразы взаимодействуют для снятия нуклеосомного барьера и удаления гистонов. Proc. Natl Acad. Sci. США 107 , 11325–11330 (2010).
Proc. Natl Acad. Sci. США 107 , 11325–11330 (2010).
Артикул PubMed Google Scholar
Джамай А., Имобердорф Р. М. и Струбин М. Непрерывный обмен гистона h3B и зависимый от транскрипции гистон h4 в дрожжевых клетках вне репликации. Мол. Ячейка 25 , 345–355 (2007).
Артикул CAS Google Scholar
Хеникофф С. Дестабилизация нуклеосом в эпигенетической регуляции экспрессии генов. Nature Rev. Genet. 9 , 15–26 (2008).
Артикул CAS PubMed Google Scholar
Инглиш, К. М., Адкинс, М. В., Карсон, Дж. Дж., Черчилль, М. Э. и Тайлер, Дж. К. Структурные основы гистоновой шаперонной активности Asf1. Cell 127 , 495–508 (2006).
Артикул CAS PubMed PubMed Central Google Scholar
Tachiwana, H. et al. Структуры нуклеосом человека, содержащие основные варианты гистона h4. Acta Crystallogr. D Biol. Кристаллогр. 67 , 578–583 (2011).
Артикул CAS PubMed Google Scholar
Эльзассер, С.J. et al. DAXX окружает димер гистона h4.3 – h5 для специфического распознавания h4.3. Природа 491 , 560–565 (2012).
Артикул CAS PubMed PubMed Central Google Scholar
Лю, К. П. и др. Структура вариантного гетеродимера гистона h4.3 – h5 в комплексе с его шапероном DAXX. Nature Struct. Мол. Биол. 19 , 1287–1292 (2012).
Артикул CAS Google Scholar
Мито, Ю., Хеникофф, Дж. Г. и Хеникофф, С. Профилирование в масштабе генома паттернов замены гистона h4.3. Nature Genet. 37 , 1090–1097 (2005).
37 , 1090–1097 (2005).
Артикул CAS PubMed Google Scholar
Wirbelauer, C., Bell, O. & Schubeler, D. Вариант гистона h4.3 откладывается в сайтах нуклеосомного смещения во всех транскрибируемых генах, в то время как активные модификации гистонов обнаруживают смещение проксимального промотора. Genes Dev. 19 , 1761–1766 (2005).
Артикул CAS PubMed PubMed Central Google Scholar
Накаяма, Т., Нисиока, К., Донг, Ю. X., Шимодзима, Т. и Хиросе, С. Drosophila Фактор GAGA направляет замену гистона h4.3, которая предотвращает распространение гетерохроматина. Genes Dev. 21 , 552–561 (2007).
Артикул CAS PubMed PubMed Central Google Scholar
Даури, Л., Chailleux, C., Bonvallet, J. & Trouche, D. Отложение гистона h4.3 в E2F-регулируемых генах связано с транскрипцией. EMBO Rep. 7 , 66–71 (2006).
Артикул CAS PubMed Google Scholar
Mito, Y., Henikoff, J. G. & Henikoff, S. Замена гистона отмечает границы цис- -регуляторных доменов. Наука 315 , 1408–1411 (2007).
Артикул CAS PubMed Google Scholar
Джин, К.и другие. Нуклеосомы, содержащие двойной вариант h4.3 / h3A.Z, маркируют «свободные от нуклеосом области» активных промоторов и других регуляторных областей. Nature Genet. 41 , 941–945 (2009). Это исследование определяет геномную локализацию хрупких двухвариантных нуклеосом.
Артикул CAS PubMed Google Scholar
Kraushaar, D. C. et al. Динамика инкорпорации по всему геному выявляет различные категории оборота для гистонового варианта h4. 3. Genome Biol. 14 , R121 (2013).
Артикул PubMed PubMed Central Google Scholar
Huang, C. et al. События расщепления тетрамера h4.3-h5 характеризуются энхансерами, специфичными для клеточного типа. PLoS Genet. 9 , e1003558 (2013).
Артикул CAS PubMed PubMed Central Google Scholar
Дил, р.Б., Хеникофф, Дж. Г. и Хеникофф, С. Полногеномная кинетика оборота нуклеосом, определяемая метаболическим мечением гистонов. Наука 328 , 1161–1164 (2010). Эта статья определяет новый метод, который использует метаболическое мечение для отслеживания обмена гистонов у многоклеточных животных.
Артикул CAS PubMed PubMed Central Google Scholar
Чоу, К. М. и др. Вариант гистона h4.3 отмечает промоторы транскрипционно активных генов во время деления клеток млекопитающих. EMBO Rep. 6 , 354–360 (2005).
Артикул CAS PubMed PubMed Central Google Scholar
Филипеску Д., Зенкер Э. и Альмоузни Г. Роль в развитии вариантов гистона h4 и их шаперонов. Trends Genet. 29 , 630–640 (2013).
Артикул CAS PubMed Google Scholar
Зенкер, Э., Ray-Gallet, D. & Almouzni, G. Двойная сторона гистонового варианта h4.3. Cell Res. 21 , 421–434 (2011).
Артикул CAS PubMed PubMed Central Google Scholar
Эльзаэссер, С. Дж., Голдберг, А. Д. и Аллис, К. Д. Новые функции для старого варианта: без замены гистона h4.3. Curr. Opin. Genet. Dev. 20 , 110–117 (2010).
Артикул CAS PubMed PubMed Central Google Scholar
Банашинский, Л. A. et al. Hira-зависимое отложение гистона h4.3 облегчает рекрутирование PRC2 в онтогенетические локусы в ES-клетках. Ячейка 155 , 107–120 (2013).
A. et al. Hira-зависимое отложение гистона h4.3 облегчает рекрутирование PRC2 в онтогенетические локусы в ES-клетках. Ячейка 155 , 107–120 (2013).
Артикул CAS PubMed Google Scholar
Суто Р. К., Кларксон М. Дж., Треметик Д. Дж. И Люгер К. Кристаллическая структура ядерной частицы нуклеосомы, содержащей вариантный гистон h3A. Z. Nature Struct. Биол. 7 , 1121–1124 (2000).
Артикул CAS Google Scholar
Meneghini, M. D., Wu, M. & Madhani, H. D. Консервированный вариант гистона h3A. Z защищает эухроматин от эктопического распространения молчащего гетерохроматина. Cell 112 , 725–736 (2003).
Артикул CAS PubMed Google Scholar
Obri, A. et al. ANP32E — это гистоновый шаперон, который удаляет h3A.Z из хроматина. Природа 505 , 648–653 (2014).
Артикул CAS PubMed Google Scholar
Smolle, M. & Workman, J. L. Модификации гистонов, связанные с транскрипцией, и криптическая транскрипция. Биохим. Биофиз. Acta 1829 , 84–97 (2013).
Артикул CAS PubMed Google Scholar
Петрук, С.и другие. Белки TrxG и PcG, но не метилированные гистоны, остаются связанными с ДНК посредством репликации. Ячейка 150 , 922–933 (2012).
Артикул CAS PubMed PubMed Central Google Scholar
Williams, S.K., Truong, D. & Tyler, J.K. Ацетилирование глобулярного ядра гистона h4 на лизине 56 способствует разборке хроматина во время активации транскрипции. Proc. Natl Acad.Sci. США 105 , 9000–9005 (2008).
Артикул PubMed Google Scholar
Tropberger, P. et al. Регуляция транскрипции посредством ацетилирования h4K122 на боковой поверхности октамера гистонов. Ячейка 152 , 859–872 (2013).
et al. Регуляция транскрипции посредством ацетилирования h4K122 на боковой поверхности октамера гистонов. Ячейка 152 , 859–872 (2013).
Артикул CAS Google Scholar
Shogren-Knaak, M. et al. Ацетилирование гистона h5K16 контролирует структуру хроматина и белковые взаимодействия. Наука 311 , 844–847 (2006).
Артикул CAS PubMed Google Scholar
Lee, J. S. et al. Пересечение гистонов между моноубиквитинированием h3B и метилированием h4, опосредованным COMPASS. Ячейка 131 , 1084–1096 (2007).
Артикул CAS PubMed Google Scholar
Venkatesh, S. et al.Set2 метилирование гистона h4 лизина 36 подавляет обмен гистонов на транскрибируемых генах. Природа 489 , 452–455 (2012).
Артикул CAS PubMed Google Scholar
Папамихос-Хронакис, М., Ватанабе, С., Рандо, О. Дж. И Петерсон, С. Л. Глобальная регуляция локализации h3A.Z ферментом ремоделирования хроматина INO80 важна для целостности генома. Cell 144 , 200–213 (2011). Эта статья характеризует роль комплекса INO80 в предотвращении неправильной локализации варианта h3A.Z в областях генома, отличных от промотора.
Артикул CAS PubMed PubMed Central Google Scholar
McKittrick, E., Gafken, P. R., Ahmad, K. & Henikoff, S. Гистон h4.3 обогащен ковалентными модификациями, связанными с активным хроматином. Proc. Natl Acad. Sci. США 101 , 1525–1530 (2004).
Артикул CAS PubMed Google Scholar
Тропбергер П. и Шнайдер Р. Почесывание (латеральной) поверхности регуляции хроматина модификациями гистонов. Nature Struct. Мол. Биол. 20 , 657–661 (2013).
Nature Struct. Мол. Биол. 20 , 657–661 (2013).
Артикул CAS Google Scholar
Тессарз П. и Кузаридес Т. Модификации гистонового ядра, регулирующие структуру и динамику нуклеосом. Nature Rev. Mol. Cell Biol. 15 , 703–708 (2014).
Артикул CAS Google Scholar
Беккер, П. Б. и Воркман, Дж. Л. Ремоделирование нуклеосом и эпигенетика. Колд Спринг Харб. Перспектива. Биол. 5 , а017905 (2013).
Артикул CAS PubMed PubMed Central Google Scholar
Кобор, М.S. et al. Белковый комплекс, содержащий консервативную Swi2 / Snf2-родственную АТФазу Swr1p, откладывает гистоновый вариант h3A.Z в эухроматин. PLoS Biol. 2 , E131 (2004).
Артикул PubMed PubMed Central Google Scholar
Watanabe, S. & Peterson, C.L. Семейство INO80 ферментов ремоделирования хроматина: регуляторы динамики вариантов гистонов. Колд Спринг Харб. Symp. Quant. Биол. 75 , 35–42 (2010).
Артикул CAS PubMed Google Scholar
Mizuguchi, G. et al. АТФ-управляемый обмен варианта гистона h3AZ, катализируемый комплексом ремоделирования хроматина SWR1. Наука 303 , 343–348 (2004).
Артикул CAS PubMed Google Scholar
Luk, E. et al. Поэтапная замена гистона на SWR1 требует двойной активации гистоном h3A.Z и каноническая нуклеосома. Ячейка 143 , 725–736 (2010).
Артикул CAS PubMed Google Scholar
Wu, W. H. et al. Swc2 представляет собой широко консервативный h3AZ-связывающий модуль, необходимый для АТФ-зависимого обмена гистонов. Nature Struct. Мол. Биол. 12 , 1064–1071 (2005).
Nature Struct. Мол. Биол. 12 , 1064–1071 (2005).
Артикул CAS Google Scholar
Ватанабе, С., Radman-Livaja, M., Rando, O.J. & Peterson, C.L. Переключатель ацетилирования гистонов регулирует отложение h3A.Z с помощью фермента ремоделирования SWRC. Наука 340 , 195–199 (2013).
Артикул CAS PubMed PubMed Central Google Scholar
Kusch, T. et al. Ацетилирование с помощью Tip60 необходимо для селективного обмена вариантами гистонов в повреждениях ДНК. Наука 306 , 2084–2087 (2004).
Артикул CAS PubMed Google Scholar
Ruhl, D. D. et al. Очистка человеческого комплекса SRCAP, который ремоделирует хроматин путем включения гистонового варианта h3A.Z в нуклеосомы. Биохимия 45 , 5671–5677 (2006).
Артикул CAS PubMed Google Scholar
Айссенберг, Дж. К., Вонг, М. и Кривиа, Дж.C. SRCAP человека и Drosophila melanogaster DOM являются гомологами, которые действуют в сигнальном пути notch. Мол. Клетка. Биол. 25 , 6559–6569 (2005).
Артикул CAS PubMed PubMed Central Google Scholar
Cai, Y. et al. Белок YL1 млекопитающих является общей субъединицей гистонацетилтрансферазы TRRAP / TIP60 и комплексов SRCAP. J. Biol. Chem. 280 , 13665–13670 (2005).
Артикул CAS PubMed Google Scholar
Johnston, H., Kneer, J., Chackalaparampil, I., Yaciuk, P. & Chrivia, J. Идентификация нового члена семейства белков SNF2 / SWI2, SRCAP, который взаимодействует с CREB-связывающим белком. J. Biol. Chem. 274 , 16370–16376 (1999).![]()
Артикул CAS PubMed Google Scholar
Конавей, Р.C. & Conaway, J. W. Комплекс ремоделирования хроматина INO80 в транскрипции, репликации и репарации. Trends Biochem. Sci. 34 , 71–77 (2009).
Артикул CAS PubMed Google Scholar
Йен, К., Винаячандран, В. и Пью, Б. Ф. Ремоделиры хроматина SWRC и INO80 распознают свободные от нуклеосом области около +1 нуклеосомы. Ячейка 154 , 1246–1256 (2013).
Артикул CAS PubMed PubMed Central Google Scholar
Д’Арси, С.и другие. Шаперон Nap1 экранирует поверхности гистонов, используемых в нуклеосомах, и может превращать h3A-h3B в нетрадиционную тетрамерную форму. Мол. Ячейка 51 , 662–677 (2013).
Артикул CAS PubMed Google Scholar
Де Конинг, Л., Корпет, А., Хабер, Дж. Э. и Альмоузни, Г. Шапероны гистонов: сеть сопровождения, регулирующая трафик гистонов. Nature Struct. Мол. Биол. 14 , 997–1007 (2007).
Артикул CAS Google Scholar
Hondele, M. et al. Структурная основа узнавания гистона h3A – h3B основным шапероном FACT. Природа 499 , 111–114 (2013).
Артикул CAS PubMed Google Scholar
Bowman, A. et al. Гистоновые шапероны Nap1 и Vps75 связывают гистоны h4 и h5 в тетрамерной конформации. Мол. Ячейка 41 , 398–408 (2011).
Артикул CAS PubMed PubMed Central Google Scholar
Сваминатан, В., Кишор, А. Х., Фебита, К. К. и Кунду, Т. К. Шаперон гистонов человека, нуклеофозмин, усиливает зависимую от ацетилирования транскрипцию хроматина. Мол. Клетка. Биол. 25 , 7534–7545 (2005).
Мол. Клетка. Биол. 25 , 7534–7545 (2005).
Артикул CAS PubMed PubMed Central Google Scholar
Лук, Э.и другие. Chz1, ядерный шаперон для гистона h3AZ. Мол. Ячейка 25 , 357–368 (2007).
Артикул CAS PubMed Google Scholar
Su, D. et al. Структурная основа распознавания h4K56-ацетилированного гистона h4 – h5 шапероном Rtt106. Природа 483 , 104–107 (2012).
Артикул CAS PubMed PubMed Central Google Scholar
Оуэн-Хьюз, Т.И Уоркман, Дж. Л. Ремоделирование структуры хроматина массива нуклеосом путем нацеленного на транскрипционный фактор замещения гистонов транс . EMBO J. 15 , 4702–4712 (1996).
Артикул CAS PubMed PubMed Central Google Scholar
Лорч Ю., Майер-Дэвис Б. и Корнберг Р. Д. Ремоделирование хроматина путем разборки нуклеосом in vitro . Proc. Natl Acad.Sci. США 103 , 3090–3093 (2006).
Артикул CAS PubMed Google Scholar
Курян Б.Г. и др. Плотность гистонов поддерживается во время транскрипции, опосредованной ремоделером хроматина RSC и гистоновым шапероном NAP1 in vitro . Proc. Natl Acad. Sci. США 109 , 1931–1936 (2012).
Артикул PubMed Google Scholar
Хан, Дж.и другие. Rtt109 ацетилирует гистон h4 лизин 56 и участвует в репликации ДНК. Наука 315 , 653–655 (2007).
Артикул CAS PubMed Google Scholar
Дрисколл, Р., Хадсон, А. и Джексон, С. П. Дрожжи Rtt109 способствуют стабильности генома за счет ацетилирования гистона h4 по лизину 56. Science 315 , 649–652 (2007).
Science 315 , 649–652 (2007).
Артикул CAS PubMed PubMed Central Google Scholar
Руфианге, А., Jacques, P.E., Bhat, W., Robert, F. & Nourani, A. Общегеномный независимый от репликации обмен гистона h4 происходит преимущественно на промоторах и включает ацетилирование h4 K56 и Asf1. Мол. Ячейка 27 , 393–405 (2007).
Артикул CAS PubMed Google Scholar
Schwabish, M. A. & Struhl, K. Asf1 опосредует вытеснение и отложение гистонов во время элонгации с помощью РНК-полимеразы II. Мол.Ячейка 22 , 415–422 (2006).
Артикул CAS PubMed Google Scholar
Колонько Е.М. и др. Каталитическая активация гистонацетилтрансферазы Rtt109 гистоновым шапероном. Proc. Natl Acad. Sci. США 107 , 20275–20280 (2010).
Артикул PubMed Google Scholar
Tsubota, T. et al. Ацетилирование гистона h4K56 катализируется гистоновыми шаперон-зависимыми комплексами. Мол. Ячейка 25 , 703–712 (2007).
Артикул CAS PubMed PubMed Central Google Scholar
Каплан Т. и др. Опосредованная клеточным циклом и шаперонами регуляция включения h4K56ac в дрожжах. PLoS Genet. 4 , e1000270 (2008).
Артикул CAS PubMed PubMed Central Google Scholar
Юделл, М.L. et al. Роль Ctk1 и Spt6 в регуляции различных состояний метилирования гистона h4 лизина 36. Mol. Клетка. Биол. 28 , 4915–4926 (2008).
Артикул CAS PubMed PubMed Central Google Scholar
Йох, С. М. , Чо, Х., Пикл, Л., Эванс, Р. М. и Джонс, К. А. Домен Spt6 Sh3 связывает Ser2P RNAPII для управления Iws1-зависимым сплайсингом и экспортом мРНК. Genes Dev. 21 , 160–174 (2007).
, Чо, Х., Пикл, Л., Эванс, Р. М. и Джонс, К. А. Домен Spt6 Sh3 связывает Ser2P RNAPII для управления Iws1-зависимым сплайсингом и экспортом мРНК. Genes Dev. 21 , 160–174 (2007).
Артикул CAS PubMed PubMed Central Google Scholar
Du, H. N., Fingerman, I. M. & Briggs, S. D. Метилирование гистона h4 K36 опосредуется путем транс- -гистонового метилирования, включающего взаимодействие между Set2 и гистоном h5. Genes Dev. 22 , 2786–2798 (2008).
Артикул CAS PubMed PubMed Central Google Scholar
Ду, Х.N. & Briggs, S. D. Поверхность нуклеосом, образованная остатками гистона h5, h3A и h4, необходима для метилирования собственно гистона h4 Lys36, ацетилирования гистона и репрессии криптической транскрипции. J. Biol. Chem. 285 , 11704–11713 (2010).
Артикул CAS PubMed PubMed Central Google Scholar
Carrozza, M. J. et al. Метилирование гистона h4 с помощью Set2 направляет деацетилирование кодирующих областей с помощью Rpd3S для подавления ложной внутригенной транскрипции. Ячейка 123 , 581–592 (2005).
Артикул CAS PubMed Google Scholar
Каплан, К. Д., Лапраде, Л. и Уинстон, Ф. Факторы элонгации транскрипции подавляют инициацию транскрипции из криптических сайтов. Наука 301 , 1096–1099 (2003).
Артикул CAS PubMed Google Scholar
Хассан, А.Х., Нили, К. Э. и Воркман, Дж. Л. Комплексы гистонацетилтрансфераз стабилизируют связывание SWI / SNF с промоторными нуклеосомами. Cell 104 , 817–827 (2001).
Артикул CAS PubMed Google Scholar
Schneiderman, J.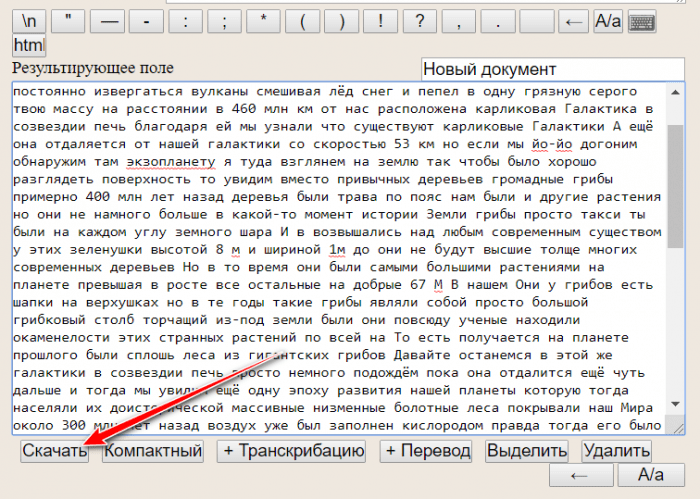 I., Orsi, G. A., Hughes, K. T., Loppin, B. & Ahmad, K. Истощенные нуклеосомами промежутки хроматина привлекают факторы сборки для варианта гистона h4.3. Proc. Natl Acad. Sci.США 109 , 19721–19726 (2012).
I., Orsi, G. A., Hughes, K. T., Loppin, B. & Ahmad, K. Истощенные нуклеосомами промежутки хроматина привлекают факторы сборки для варианта гистона h4.3. Proc. Natl Acad. Sci.США 109 , 19721–19726 (2012).
Артикул PubMed Google Scholar
Дрю, Х. Р. и Трэверс, А. А. Изгиб ДНК и его связь с позиционированием нуклеосом. J. Mol. Биол. 186 , 773–790 (1985).
Артикул CAS PubMed Google Scholar
Segal, E. et al. Геномный код для позиционирования нуклеосом. Природа 442 , 772–778 (2006).
Артикул CAS PubMed PubMed Central Google Scholar
Hartley, P. D. & Madhani, H. D. Механизмы, определяющие расположение и идентичность промоторных нуклеосом. Cell 137 , 445–458 (2009).
Артикул CAS PubMed PubMed Central Google Scholar
Гангули, Д., Chereji, R. V., Iben, J. R., Cole, H. A. & Clark, D. J. RSC-зависимая конструктивная и деструктивная интерференция между противостоящими массивами фазированных нуклеосом в дрожжах. Genome Res. 24 , 1637–1649 (2014).
Артикул CAS PubMed PubMed Central Google Scholar
Ranjan, A. et al. Область, свободная от нуклеосом, доминирует над ацетилированием гистонов при нацеливании SWR1 на промоторы для замены h3A.Z. Ячейка 154 , 1232–1245 (2013). Ссылки 54–56 и 94 определяют факторы, регулирующие функцию SWR при замене гистонового варианта h3A.Z.
Артикул CAS PubMed Google Scholar
Юань, G.C. et al. Геномная идентификация положений нуклеосом в S. cerevisiae . Наука 309 , 626–630 (2005).
Артикул CAS PubMed Google Scholar
Маврич Т.N. et al. Модель барьерной нуклеосомы для статистического позиционирования нуклеосом по всему геному дрожжей. Genome Res. 18 , 1073–1083 (2008).
Артикул CAS PubMed PubMed Central Google Scholar
Zofall, M. et al. Гистон h3A.Z взаимодействует с факторами гетерохроматина РНКи, подавляя антисмысловые РНК. Природа 461 , 419–422 (2009).
Артикул CAS PubMed PubMed Central Google Scholar
Чжан, Х., Робертс, Д. Н. и Кэрнс, Б. Р. Полногеномная динамика Htz1, варианта гистона h3A, который настраивает репрессированные / базальные промоторы для активации через потерю гистонов. Cell 123 , 219–231 (2005).
Артикул CAS PubMed PubMed Central Google Scholar
Dion, M. F. et al. Динамика независимого от репликации оборота гистонов у почкующихся дрожжей. Наука 315 , 1405–1408 (2007). Ссылки 77 и 99 определяют концепцию, геномное распределение и факторы, управляющие обменом гистонов у дрожжей дикого типа.
Артикул CAS PubMed Google Scholar
Durant, M. & Pugh, B. F. NuA4-направленные транзакции хроматина по всему геному Saccharomyces cerevisiae . Мол. Клетка. Биол. 27 , 5327–5335 (2007).
Артикул CAS PubMed PubMed Central Google Scholar
Альтаф, М.и другие. NuA4-зависимое ацетилирование нуклеосомных гистонов h5 и h3A напрямую стимулирует включение h3A. Z комплексом SWR1. J. Biol. Chem. 285 , 15966–15977 (2010).
Артикул CAS PubMed PubMed Central Google Scholar
Галлей, Дж.![]() Э., Каплан, Т., Ван, А. Ю., Кобор, М. С. и Райн, Дж. Роли для h3A. Z и его ацетилирование в транскрипции GAL1 и индукции гена, но не в транскрипционной памяти GAL1. PLoS Biol. 8 , e1000401 (2010).
Э., Каплан, Т., Ван, А. Ю., Кобор, М. С. и Райн, Дж. Роли для h3A. Z и его ацетилирование в транскрипции GAL1 и индукции гена, но не в транскрипционной памяти GAL1. PLoS Biol. 8 , e1000401 (2010).
Артикул CAS PubMed PubMed Central Google Scholar
Draker, R. et al. Комбинация ацетилирования h3A.Z и h5 рекрутирует Brd2 на хроматин во время активации транскрипции. PLoS Genet. 8 , e1003047 (2012).
Артикул CAS PubMed PubMed Central Google Scholar
Чен, П.и другие. h4.3 активно маркирует энхансеры и запускает транскрипцию генов, открывая хроматин более высокого порядка. Genes Dev. 27 , 2109–2124 (2013).
Артикул CAS PubMed PubMed Central Google Scholar
Heinz, S. et al. Выбор и функция энхансеров, специфичных для определенного типа клеток. Nature Rev. Mol. Cell Biol. http://dx.doi.org/10.1038/nrm3949 (2015).
Конерли, М.L. et al. Изменения в занятости h3A.Z и метилировании ДНК во время В-клеточного лимфомагенеза. Genome Res. 20 , 1383–1390 (2010).
Артикул CAS PubMed PubMed Central Google Scholar
Zilberman, D., Coleman-Derr, D., Ballinger, T. & Henikoff, S. Гистон h3A.Z и метилирование ДНК являются взаимно антагонистическими метками хроматина. Природа 456 , 125–129 (2008).
Артикул CAS PubMed PubMed Central Google Scholar
Пчелинцев Н.A. et al. Размещение гистонового шаперонного комплекса HIRA в хроматиновом ландшафте. Cell Rep. 3 , 1012–1019 (2013).
Артикул CAS PubMed PubMed Central Google Scholar
Киреева М. Л. и др. Ремоделирование нуклеосом, индуцированное РНК-полимеразой II: потеря димера h3A / h3B во время транскрипции. Мол. Ячейка 9 , 541–552 (2002).
Л. и др. Ремоделирование нуклеосом, индуцированное РНК-полимеразой II: потеря димера h3A / h3B во время транскрипции. Мол. Ячейка 9 , 541–552 (2002).
Артикул CAS PubMed Google Scholar
Кулаева, О.I. et al. Механизм ремоделирования и восстановления хроматина при пассаже РНК-полимеразы II. Nature Struct. Мол. Биол. 16 , 1272–1278 (2009).
Артикул CAS Google Scholar
Белоцерковская Р. и др. FACT способствует зависимому от транскрипции изменению нуклеосом. Наука 301 , 1090–1093 (2003).
Артикул CAS PubMed Google Scholar
Паври, Р.и другие. Моноубиквитинирование гистона h3B действует совместно с FACT, регулируя удлинение с помощью РНК-полимеразы II. Cell 125 , 703–717 (2006).
Артикул CAS PubMed Google Scholar
Hsieh, F. K. et al. Действие гистоновых шаперонов FACT при транскрипции через хроматин РНК-полимеразой II. Proc. Natl Acad. Sci. США 110 , 7654–7659 (2013).
Артикул PubMed Google Scholar
Сарай, Н.и другие. WHSC1 связывает удлинение транскрипции с HIRA-опосредованным отложением гистона h4.3. EMBO J. 32 , 2392–2406 (2013).
Артикул CAS PubMed PubMed Central Google Scholar
Кулаева О.И., Сие Ф.К., Чанг Х.В., Лузе Д.С. и Студицкий В.М. Механизм транскрипции через нуклеосому с помощью РНК-полимеразы II. Биохим. Биофиз. Acta 1829 , 76–83 (2013).
Артикул CAS PubMed Google Scholar
Fleming, A. B., Kao, C.![]() F., Hillyer, C., Pikaart, M. & Osley, M. A. Убиквитилирование h3B играет роль в динамике нуклеосом во время элонгации транскрипции. Мол. Ячейка 31 , 57–66 (2008).
F., Hillyer, C., Pikaart, M. & Osley, M. A. Убиквитилирование h3B играет роль в динамике нуклеосом во время элонгации транскрипции. Мол. Ячейка 31 , 57–66 (2008).
Артикул CAS PubMed Google Scholar
Джоши, А.А. и Струл, К.Взаимодействие хромодомена Eaf3 с метилированным h4K36 связывает деацетилирование гистонов с элонгацией Pol II. Мол. Ячейка 20 , 971–978 (2005).
Артикул CAS PubMed Google Scholar
Cheung, V. et al. Факторы, связанные с хроматином и транскрипцией, подавляют транскрипцию внутри кодирующих областей по всему геному Saccharomyces cerevisiae . PLoS Biol. 6 , e277 (2008). Это исследование подчеркивает роль хроматина в регуляции транскрипции с криптических промоторов.
Артикул CAS PubMed PubMed Central Google Scholar
Gossett, A. J. & Lieb, J. D. In vivo эффекты истощения гистона h4 на занятость и положение нуклеосом в Saccharomyces cerevisiae . PLoS Genet. 8 , e1002771 (2012).
Артикул CAS PubMed PubMed Central Google Scholar
Йох, С.М., Лукас, Дж. С. и Джонс, К. А. Комплекс Iws1: Spt6: CTD контролирует котранскрипционный биосинтез мРНК и опосредованное HYPB / Setd2 метилирование гистона h4K36. Genes Dev. 22 , 3422–3434 (2008).
Артикул CAS PubMed PubMed Central Google Scholar
Smolle, M. et al. Ремоделирующие хроматин Isw1 и Chd1 поддерживают структуру хроматина во время транскрипции, предотвращая обмен гистонов. Nature Struct. Мол. Биол. 19 , 884–892 (2012). Ссылки 46 и 121 определяют механизм, участвующий в восстановлении хроматина после транскрипции.
Артикул CAS Google Scholar
Винклер, Д. Д., Мутураджан, У. М., Хиб, А. Р. и Люгер, К. Шаперон гистонов FACT координирует взаимодействие нуклеосом посредством множественных синергетических событий связывания. J. Biol. Chem. 286 , 41883–41892 (2011).
Артикул CAS PubMed PubMed Central Google Scholar
Carvalho, S. et al. Гистон-метилтрансфераза SETD2 координирует набор FACT с динамикой нуклеосом во время транскрипции. Nucleic Acids Res. 41 , 2881–2893 (2013).
Артикул CAS PubMed PubMed Central Google Scholar
Мальтби, В.E. et al. Метилирование гистона h4 лизина 36 нацелено на ремоделирующий комплекс Isw1b на хроматин. Мол. Клетка. Биол. 32 , 3479–3485 (2012).
Артикул CAS PubMed PubMed Central Google Scholar
Radman-Livaja, M. et al. Ключевая роль Chd1 в динамике гистона h4 на 3′-концах длинных генов у дрожжей. PLoS Genet. 8 , e1002811 (2012).
Артикул CAS PubMed PubMed Central Google Scholar
Хенниг, Б.П., Бендрин, К., Чжоу, Ю. и Фишер, Т. Ремоделеры хроматина Chd1 поддерживают организацию нуклеосом и подавляют криптическую транскрипцию. EMBO Rep. 13 , 997–1003 (2012).
Артикул CAS PubMed PubMed Central Google Scholar
Lee, J. S. et al. Созависимость моноубиквитинирования h3B и повторной сборки нуклеосом на Chd1. Genes Dev. 26 , 914–919 (2012).
Артикул CAS PubMed PubMed Central Google Scholar
Батта, К. , Zhang, Z., Yen, K., Goffman, D. B. & Pugh, B. F. Полногеномная функция убиквитилирования h3B в промоторных и генных областях. Genes Dev. 25 , 2254–2265 (2011).
, Zhang, Z., Yen, K., Goffman, D. B. & Pugh, B. F. Полногеномная функция убиквитилирования h3B в промоторных и генных областях. Genes Dev. 25 , 2254–2265 (2011).
Артикул CAS PubMed PubMed Central Google Scholar
Говинд, К. К. и др. Фосфорилированный Pol II CTD привлекает несколько HDAC, включая Rpd3C (S), для зависимого от метилирования деацетилирования нуклеосом ORF. Мол.Ячейка 39 , 234–246 (2010).
Артикул CAS PubMed PubMed Central Google Scholar
Goldberg, A. D. et al. Определенные факторы контролируют локализацию варианта гистона h4.3 в определенных областях генома. Cell 140 , 678–691 (2010).
Артикул CAS PubMed PubMed Central Google Scholar
Оои, С.Л., Присс, Дж. Р. и Хеникофф, С. Динамика варианта гистона h4.3 в зародышевой линии Caenorhabditis elegans . PLoS Genet. 2 , e97 (2006).
Артикул CAS PubMed PubMed Central Google Scholar
Loyola, A. & Almouzni, G. Маркировка вариантов гистона h4: как, когда и почему? Trends Biochem. Sci. 32 , 425–433 (2007).
Артикул CAS PubMed Google Scholar
Вэнь, Х.и другие. ZMYND11 связывает гистон h4.3K36me3 с удлинением транскрипции и подавлением опухоли. Природа 508 , 263–268 (2014).
Артикул CAS PubMed PubMed Central Google Scholar
Guo, R. et al. BS69 / ZMYND11 считывает и связывает хроматин, декорированный триметилированием гистона h4.3 лизина 36, с регулируемым процессингом пре-мРНК. Мол. Ячейка 56 , 298–310 (2014).![]() Ссылки 133 и 134 определяют идентификацию человеческого белка, ZMYND11, который специфически распознает вариант h4 метилированного гистона h4K36.3, и это участвует в элонгации Pol II и сплайсинге РНК.
Ссылки 133 и 134 определяют идентификацию человеческого белка, ZMYND11, который специфически распознает вариант h4 метилированного гистона h4K36.3, и это участвует в элонгации Pol II и сплайсинге РНК.
Артикул CAS PubMed PubMed Central Google Scholar
Wen, H., Li, Y., Li, H. & Shi, X. ZMYND11: специфичный для h4.3 считыватель h4K36me3. Cell Cycle 13 , 2153–2154 (2014).
Артикул CAS PubMed Google Scholar
Елинич П., Pellegrino, J. & David, G. Новый комплекс млекопитающих, содержащий Sin3B, смягчает ацетилирование гистонов и прогрессирование РНК-полимеразы II в транскрибируемых локусах. Мол. Клетка. Биол. 31 , 54–62 (2011).
Артикул CAS PubMed Google Scholar
Kumar, G. S. et al. Требования к последовательности для комбинаторного распознавания гистона h4 субъединицами MRG15 и Pf1 корепрессорного комплекса Rpd3S / Sin3S. J. Mol. Биол. 422 , 519–531 (2012).
Артикул CAS PubMed PubMed Central Google Scholar
Li, B. et al. Совместное действие PHD и хромодоменов направляет Rpd3S HDAC на транскрибируемый хроматин. Наука 316 , 1050–1054 (2007).
Артикул CAS PubMed Google Scholar
Эстеллер, М.Некодирующие РНК при заболеваниях человека. Nature Rev. Genet. 12 , 861–874 (2011).
Артикул CAS PubMed Google Scholar
Зовойлис А. и др. Уровень экспрессии малых некодирующих РНК, полученных из первого экзона генов, кодирующих белок, позволяет прогнозировать статус рака. EMBO Rep. 15 , 402–410 (2014).
Артикул CAS PubMed PubMed Central Google Scholar
Цзян, Л.и другие. PfSETvs метилирование гистона h4K36 репрессирует гены вирулентности в Plasmodium falciparum . Природа 499 , 223–227 (2013).
Артикул CAS PubMed PubMed Central Google Scholar
Луко, Р. Ф. и др. Регулирование альтернативного сплайсинга модификациями гистонов. Наука 327 , 996–1000 (2010).
Артикул CAS PubMed PubMed Central Google Scholar
Прадипа, М.М., Сазерленд, Х. Г., Уле, Дж., Граймс, Г. Р. и Бикмор, В. А. Psip1 / Ledgf p52 связывает метилированный гистон h4K36 и факторы сплайсинга и способствует регуляции альтернативного сплайсинга. PLoS Genet. 8 , e1002717 (2012). Ссылки 142 и 143 перечисляют роль структуры и модификации хроматина в котранскрипционной регуляции альтернативного сплайсинга.
Артикул CAS PubMed PubMed Central Google Scholar
Луко, Р.Ф., Алло, М., Шор, И. Е., Корнблихтт, А. Р. и Мистели, Т. Эпигенетика в альтернативном сплайсинге пре-мРНК. Cell 144 , 16–26 (2011).
Артикул CAS PubMed PubMed Central Google Scholar
Kolasinska-Zwierz, P. et al. Дифференциальное маркирование хроматина интронов и экспрессируемых экзонов с помощью h4K36me3. Nature Genet. 41 , 376–381 (2009).
Артикул CAS PubMed Google Scholar
Бартоломей, М.С., Халден, Н. Ф., Каллен, К. Р. и Корден, Дж. Л. Генетический анализ повторяющегося карбоксиконцевого домена самой большой субъединицы РНК-полимеразы II мыши. Мол. Клетка. Биол. 8 , 330–339 (1988).
Биол. 8 , 330–339 (1988).
Артикул CAS PubMed PubMed Central Google Scholar
Litingtung, Y. et al. Задержка роста и неонатальная летальность у мышей с гомозиготной делецией в С-концевом домене РНК-полимеразы II. Мол. Genet Genet. 261 , 100–105 (1999).
Артикул CAS PubMed Google Scholar
Hsin, J. P. & Manley, J. L. CTD РНК-полимеразы II координирует транскрипцию и процессинг РНК. Genes Dev. 26 , 2119–2137 (2012).
Артикул CAS PubMed PubMed Central Google Scholar
Хайдеманн, М., Hintermair, C., Voss, K. & Eick, D. Динамические паттерны фосфорилирования CTD РНК-полимеразы II во время транскрипции. Биохим. Биофиз. Acta 1829 , 55–62 (2013).
Артикул CAS PubMed Google Scholar
Смолле М. и Венкатеш С. в книге Основы хроматина (ред. Воркман, Дж. Л. и Абмайр, С. М.) (Springer, 2014).
Google Scholar
Дил, р.Б. и Хеникофф, С. Захват динамического эпигенома. Genome Biol. 11 , 218 (2010).
Артикул CAS PubMed PubMed Central Google Scholar
Кимура, Х. и Кук, П. Р. Кинетика ядер гистонов в живых клетках человека: небольшой обмен h4 и h5 и некоторый быстрый обмен h3B. J. Cell Biol. 153 , 1341–1353 (2001).
Артикул CAS PubMed PubMed Central Google Scholar
Восс, Т.К. и Хагер, Г. Л. Визуализация динамики хроматина в интактных клетках. Биохим. Биофиз. Acta 1783 , 2044–2051 (2008).
Артикул CAS PubMed PubMed Central Google Scholar
Misteli, T. , Gunjan, A., Hock, R., Bustin, M. & Brown, D. T. Динамическое связывание гистона h2 с хроматином в живых клетках. Nature 408 , 877–881 (2000).
, Gunjan, A., Hock, R., Bustin, M. & Brown, D. T. Динамическое связывание гистона h2 с хроматином в живых клетках. Nature 408 , 877–881 (2000).
Артикул CAS PubMed Google Scholar
Ray-Gallet, D.и другие. Динамика отложения гистона h4 in vivo выявляет механизм заполнения нуклеосомных пробелов для h4.3, чтобы поддерживать целостность хроматина. Мол. Ячейка 44 , 928–941 (2011).
Артикул CAS PubMed Google Scholar
Verzijlbergen, K. F. et al. Индуцированный рекомбинацией обмен тегами для отслеживания старых и новых белков. Proc. Natl Acad. Sci. США 107 , 64–68 (2010).
Артикул PubMed Google Scholar
Xu, M. et al. Разделение тетрамеров гистонов h4 – h5 во время сборки хроматина, зависимой от репликации ДНК. Наука 328 , 94–98 (2010).
Артикул CAS PubMed Google Scholar
Свит, С. М., Ли, М., Томас, П. М., Дурбин, К. Р. и Келлехер, Н. Л. Кинетика восстановления меток метилирования h4K79 в глобальном хроматине человека. J. Biol. Chem. 285 , 32778–32786 (2010).
Артикул CAS PubMed PubMed Central Google Scholar
Venkatesh, S. & Workman, J. L. Распознавание варианта метилированного гистона h4.3 для предотвращения опухолей. Cell Res. 24 , 649–650 (2014).
Артикул CAS PubMed PubMed Central Google Scholar
Канг, Б.и другие. Фосфорилирование h5 Ser 47 способствует HIRA-опосредованной сборке нуклеосом. Genes Dev. 25 , 1359–1364 (2011).
Артикул CAS PubMed PubMed Central Google Scholar
Straube, K. , Blackwell, J. S. Jr & Pemberton, L. F. Nap1 и Chz1 имеют отдельные функции импорта и сборки ядер Htz1. Трафик 11 , 185–197 (2010).
, Blackwell, J. S. Jr & Pemberton, L. F. Nap1 и Chz1 имеют отдельные функции импорта и сборки ядер Htz1. Трафик 11 , 185–197 (2010).
Артикул CAS PubMed Google Scholar
Имбо, Д., Gamar, L., Rufiange, A., Paquet, E. & Nourani, A. Гистоновый шаперон Rtt106 функционально связан с элонгацией транскрипции и участвует в регуляции ложной транскрипции с криптических промоторов в дрожжах. J. Biol. Chem. 283 , 27350–27354 (2008).
Артикул CAS PubMed Google Scholar
Мао, З. и др. Anp32e, гистоновый шаперон высших эукариот, направляет преимущественное распознавание h3A. Z. Cell Res. 24 , 389–399 (2014).
Артикул CAS Google Scholar
AP2 транскрипционный фактор CBX1 со специфической функцией в симбиотическом обмене питательными веществами у микоризного лотоса японского
Значение
Арбускулярные микоризные (AM) грибы способствуют поглощению фосфора растениями-хозяевами в обмен на органический углерод. Эксперименты с физиологическими индикаторами показали, что до 100% приобретенного фосфата может доставляться растениям через путь поглощения микоризного фосфата (MPU).Предыдущие исследования показали, что CTTC cis -регуляторный элемент (CRE) необходим для активации промотора микоризы-специфического переносчика фосфата и генов H + -АТФазы. Однако точный механизм транскрипции, непосредственно контролирующий MPU, неизвестен. Здесь мы показываем, что CBX1 связывает CTTC и AW-box CRE и координирует микоризный переносчик фосфата и гены H + -АТФазы. Интересно, что гены, участвующие в биосинтезе липидов, также регулируются CBX1 посредством связывания с AW box, включая RAM2 .Наша работа предлагает общий регуляторный механизм, лежащий в основе комплексного контроля характеристик симбиотического обмена питательными веществами.
Abstract
Симбиоз арбускулярного микориза (AM), широко распространенная мутуалистическая ассоциация между наземными растениями и грибами, зависит от взаимного обмена фосфором, вызванного поглощением протон-связанного фосфата растениями-хозяевами и углерода, поставляемого AM-грибам зависимым от хозяина сахаром и биосинтез липидов. Молекулярные механизмы и модули регуляции цис-, лежащие в основе контроля поглощения фосфата и синтеза жирных кислот de novo в AM-симбиозе, плохо изучены.Здесь мы показываем, что фактор транскрипции семейства AP2 CTTC MOTIF-BINDING TRANSCRIPTION FACTOR1 (CBX1), гомолог WRINKLED1 (WRI1), напрямую связывает эволюционно консервативный мотив CTTC, который обогащен генами, регулируемыми микоризой, и активирует Lotus japonicus transporter pho1963 4 ( LjPT4 ) in vivo и in vitro. Более того, индуцируемый микоризой ген, кодирующий H + -АТФазу ( LjHA1 ), участвующий в активизации поглощения питательных веществ на симбиотическом интерфейсе через периарскулярную мембрану, ко-регулируется с LjPT4 посредством CBX1.Соответственно, мутанты с дефектом CBX1 и демонстрируют пониженную микоризную колонизацию. Более того, профили связывания по всему геному, исследования связывания ДНК и гетерологичная экспрессия выявляют дополнительное связывание CBX1 с AW box, консенсусным ДНК-связывающим мотивом для WRI1, который обогащен промоторами генов гликолиза и биосинтеза жирных кислот. Мы показываем, что CBX1 активирует экспрессию липидных метаболических генов, включая глицерин-3-фосфатацилтрансферазу RAM2 , участвующих в биосинтезе ацилглицерина.Наше открытие определяет роль CBX1 как регулятора генов-хозяев, участвующих в захвате фосфата и синтезе липидов посредством связывания с молекулярным модулем CTTC / AW, и поддерживает модель, лежащую в основе двунаправленного обмена фосфором и углеродом, фундаментальной черты мутуалистического симбиоза AM. .
Симбиоз арбускулярной микоризы (AM) — это тесная ассоциация между грибами типа Glomeromycota и корнями наземных растений, которые совместно эволюционировали более 400 млн лет назад (1). Характерным эффектом AM-симбиоза является усиление поглощения фосфора в форме неорганического фосфата (Pi) от AM-грибов растением-хозяином в обмен на фотосинтетически закрепленный углерод (2, 3). После проникновения в корковые клетки гифы грибов образуют дихотомически разветвленные арбускулы, окруженные периарбускулярной мембраной растений (PAM), которая служит интерфейсом для обмена питательными веществами между симбионтами. Индуцируемые микоризой переносчики Pi находятся в PAM (4⇓ – 6) и необходимы для функции и поддержания арбускул.Дефектные аллели Medicago truncatula MtPT4 , OsPT11 риса и ZmPT6 кукурузы ZmPT6 сильно нарушали путь поглощения микоризного фосфата (MPU) и ускоряли дегенерацию арбускул (7–9). Индуцируемые микоризой переносчики Pi принадлежат к подсемействам I, II и III семейства переносчиков Pi растений 1 (Pht1), которые примерно сгруппированы в четыре подсемейства (10-12). Подсемейство I содержит переносчики Pi, экспрессируемые исключительно в микоризных корнях; некоторые представители подсемейства II и III индуцируются микоризой; подсемейство IV состоит из переносчиков Pi из однодольных, которые не индуцируются микоризой.Анализ последовательной делеции промоторных элементов продемонстрировал регуляторную роль CTTC CRE (CTTCTTGTTC, альтернативно названного «MYCS», TTTCTTGTTCT) в генах переносчиков Pi, индуцируемых микоризой (13–16).
Характерным эффектом AM-симбиоза является усиление поглощения фосфора в форме неорганического фосфата (Pi) от AM-грибов растением-хозяином в обмен на фотосинтетически закрепленный углерод (2, 3). После проникновения в корковые клетки гифы грибов образуют дихотомически разветвленные арбускулы, окруженные периарбускулярной мембраной растений (PAM), которая служит интерфейсом для обмена питательными веществами между симбионтами. Индуцируемые микоризой переносчики Pi находятся в PAM (4⇓ – 6) и необходимы для функции и поддержания арбускул.Дефектные аллели Medicago truncatula MtPT4 , OsPT11 риса и ZmPT6 кукурузы ZmPT6 сильно нарушали путь поглощения микоризного фосфата (MPU) и ускоряли дегенерацию арбускул (7–9). Индуцируемые микоризой переносчики Pi принадлежат к подсемействам I, II и III семейства переносчиков Pi растений 1 (Pht1), которые примерно сгруппированы в четыре подсемейства (10-12). Подсемейство I содержит переносчики Pi, экспрессируемые исключительно в микоризных корнях; некоторые представители подсемейства II и III индуцируются микоризой; подсемейство IV состоит из переносчиков Pi из однодольных, которые не индуцируются микоризой.Анализ последовательной делеции промоторных элементов продемонстрировал регуляторную роль CTTC CRE (CTTCTTGTTC, альтернативно названного «MYCS», TTTCTTGTTCT) в генах переносчиков Pi, индуцируемых микоризой (13–16).
Движущей силой притока клеточного Pi является протонный градиент, генерируемый H + -АТФазой, которая активирует симпорт H + / Pi через транспортеры Pht1 в плазматической мембране (17). В M. truncatula и рис, индуцируемая микоризой H + -АТФаза (HA1) важна для развития MPU и arbuscule (18, 19).Более того, регуляторная роль CTTC CRE в промоторе индуцируемого микоризой гена H + -АТФазы SlHA8 была продемонстрирована на томате (20). Хотя CTTC CRE широко присутствует в генах, чувствительных к микоризе, факторы транскрипции, нацеленные на CTTC CRE, остаются неуловимыми.
AP2 относятся к суперсемейству AP2 / ERF и подразделяются на подсемейства, подобные WRINKLED1, APETALA и AINTEGUMENTA (21, 22). В Arabidopsis thaliana WRINKLED1 (WRI1) регулирует гены, кодирующие ферменты позднего гликолиза и биосинтеза жирных кислот посредством связывания с боксом AW [CnTnG (n) 7 CG] во время созревания семян, при этом требуются WRI1, WRI3 и WRI4 для биосинтеза кутина в тканях цветка (22–25).В симбиозе AM растения обеспечивают микоризные грибы углеводами и жирными кислотами в качестве источника углерода для поддержания мутуализма (26–31). Микоризные специфические для хозяина гены WRI в M. truncatula были обозначены как MtWRI5a / b / c . Подобно A. thaliana WRI1 , сверхэкспрессия MtWRI5a / b / c приводит к накоплению триацилглицерина (ТАГ) в листьях табака (29). Соответственно, искусственное замалчивание микроРНК MtWRI5b приводило к более низкому уровню микоризной колонизации (32).Здесь мы идентифицируем индуцируемый микоризой WRI1-подобный AP2 фактор транскрипции CBX1, который одновременно регулирует центральные компоненты MPU и биосинтез микоризных липидов посредством прямого связывания с CTTC и мотивами AW-бокса в промоторах целевых генов. Мы предполагаем, что CBX1, вероятно, играет центральную роль в эволюции и поддержании AM симбиоза.
Результаты
CBX1 кодирует фактор транскрипции, содержащий домен AP2, который связывается с CTTC cis -Действующим регуляторным элементом. Чтобы изучить функцию мотива CTTC в промоторе LjPT4 , химерные конструкции промотора LjPT4 с репортерным геном β-глюкуронидазы ( pLjPT4: GUS ), содержащим мотив CTTC или его мутантную форму (mCTTC) были стабильно трансформированы в корни Lotus japonicus (рис. 1 A ). Промотор LjPT4 и четырехкратный тандемный повтор CTTCTTGTTC, слитый с минимальным промотором вируса мозаики цветной капусты 35S (4 * CTTC), специфически направляли активность GUS в корнях, содержащих арбускулы, подтверждая предыдущие результаты (14, 33), в то время как присутствие mCTTC привело к значительному снижению активности промотора LjPT4 (рис. 1 A и SI Приложение , рис. S1 A и B ). Иногда остаточная pLjPT4 — экспрессия mCTTC: GUS обнаруживалась и ограничивалась арбускул-содержащими клетками ( SI Приложение , рис. S1 C ), что указывает на действие альтернативных цис- элементов в транскрипционной активации . Выражение LjPT4 . Таким образом, мы продемонстрировали, что мотив CTTC необходим, но недостаточен для полной активности промотора LjPT4 в микоризных секторах корня (рис.1 A и SI Приложение , рис. S1 C ).
Последовательно-специфические ДНК-связывающие свойства CBX1. ( A ) CTTC требуется для регуляции гена LjPT4 в микоризных корнях. На схематической диаграмме показаны pLjPT4: GUS и pLjPT4-mCTTC: GUS с мутациями ( Upper ). Нижний показывает активность GUS в трансгенных волосатых корнях, несущих различные репортеры, в присутствии R. irregularis .EV, вектор pRedRoot-GUS; 4 * CTTC, четырехкратный тандемный повтор CTTCTTGTTC, слитый с минимальным 35S промотором вируса мозаики цветной капусты; 4-MU, 4-метилумбеллиферон. Среднее ± стандартное отклонение ( n = 3). Использовался критерий Краскела – Уоллиса с последующим критерием наименьшего значимого различия Фишера [Краскал – Уоллис χ 2 = 9,7, степень свободного (df) = 3, P <0,05]. Было проведено три независимых эксперимента с аналогичными результатами. ( B ) EMSA связывания CBX1 с мотивом CTTC. Немеченый CTTC CRE или mCTTC CRE использовали в качестве конкурента.Сверху указано возрастающее количество ДНК конкурента. Красная стрелка указывает на комплекс белок-ДНК. ( C ) Логотип последовательности CTTC CRE был создан из 21 предполагаемого мотива CTTC в промоторах 19 генов переносчиков Pi, индуцируемых микоризой, показанных в Приложении SI , Таблица S2 с использованием WebLogo (weblogo. berkeley.edu/logo.cgi) . Высота стопки представляет собой степень сохранения, а размер букв представляет относительную частоту. ( D ) Предпочтение связывания ДНК CBX1 с мотивом CTTC в EMSA.Были синтезированы девять меченых Cy5 олигонуклеотидов, несущих замены одной пары оснований. WT, мотив CTTC дикого типа; красные буквы, базовые изменения в CTTC; черные буквы, базы дикого типа. ( E ) Схематическая диаграмма усеченных белков CBX1. AP2, домен APETALA2; NLS, сигнал ядерной локализации. Слева обозначены белковые области. ( F ) Относительная аффинность связывания укороченного CBX1 на мотиве CTTC (w) или мутированного CTTC (m) в EMSA.
Чтобы идентифицировать факторы транскрипции, которые напрямую связываются с промотором LjPT4 , гены-кандидаты, которые реагировали на AM-грибы Rhizophagus irregularis и Gigaspora margarita (34, 35), были отобраны для исследований связывания белок-ДНК с использованием электрофоретического метода. анализ сдвига подвижности (EMSA).Мы обнаружили, что белок, кодируемый геном Lj6g3v1048880.1, опосредует отчетливый сдвиг промоторной ДНК и мотива CTTC, соответственно ( SI, приложение , рис. S2 A и B ). Связывание вытеснялось немеченым CTTC, но не mCTTC (фиг. 1 B ). Исследуемый белок в дальнейшем именуется CTTC-BINDING TRANSCRIPTION FACTOR1 (CBX1). CBX1 — это фактор транскрипции семейства AP2, который принадлежит 15 членам семейства AP2 L. japonicus с типичными двойными доменами AP2 (21).AM-грибки активировали пять факторов транскрипции AP2, в том числе CBX1 , WRI5a (Lj2g3v1338890.1 и Lj2g3v1338880.1), WRI5b (Lj1g3v2952280.1 и Lj1g3v2952280.1 и Lj1g10c21962.192.1) ) и WRI3 (Lj0g3v0151469.1), которые все сгруппированы с WRI1-подобным подсемейством ( SI Приложение , рис. S3) (34, 36). WRI5a , WRI5b, и WRI5c были индуцированы сверхэкспрессией CBX1 в L. japonicus , и их кодирующие белки могут слабо связывать CTTC-содержащую ДНК in vitro ( SI Приложение , рис. S2 A — C ).
japonicus , и их кодирующие белки могут слабо связывать CTTC-содержащую ДНК in vitro ( SI Приложение , рис. S2 A — C ).
Для картирования взаимодействия мотивов CBX1-CTTC при разрешении одного нуклеотида, для EMSA были разработаны замены одной пары оснований в мотиве CTTC (CTTCTGTTC) (рис. 1 C и SI, приложение , таблица S1). Первый C в мотиве CTTC не был мутирован в синтетических олигонуклеотидах, поскольку он не консервативен в MYCS ( SI, приложение , таблица S2).EMSA показал сильно сниженную аффинность связывания CBX1 с олигомерами CTTC с изменениями оснований в положениях T 3 , T 5 , T 6 или G 7 , тогда как изменения только в C 4 или T 8 умеренно влияет на связывание ДНК (фиг. 1 D ). Таким образом, наши данные показали, что TCTTGT является основным мотивом, удовлетворяющим минимальным требованиям к последовательности для связывания ДНК с высоким сродством CBX1. Чтобы определить белковые участки CBX1, ответственные за связывание ДНК, были созданы различные формы усеченного CBX1 для исследований связывания ДНК (рис.1 E ). Два домена AP2 не смогли связать элемент CTTC. Сравнение связывающей способности CBX 1–308 и CBX 41–308 выявило ограниченный эффект N-конца, охватывающего 40 аминокислот. Присутствие перекрывающих домен аминокислот 212–308 в комбинации с доменами AP2 делает возможным связывание ДНК (рис. 1 F ). В C-концевой части CBX1 важную роль в связывании CTTC можно отнести к области, охватывающей аминокислоты 271-308.
CBX1 необходим для правильной колонизации микоризных корней.
Чтобы исследовать функцию CBX1, два мутанта cbx1-2 и cbx1-3 , несущие вставки LORE1a в последнем экзоне или в 5′-UTR ( SI Приложение , рис. S4 A и B) ) были исследованы на микоризные фенотипы, выращенные при низком уровне Pi (100 мкМ) в присутствии или в отсутствие R. irregularis (37) (рис. 2 A ). Сильно сниженная колонизация [общая (%)] наблюдалась в обеих мутантных линиях по сравнению с диким типом через 6 недель после инокуляции (рис.2 А ). Кроме того, доля секторов корня, содержащих арбускулы гриба {[A + V + H (%)] и [A + H (%)]}, была значительно ниже в мутантных линиях, чем в линиях дикого типа (рис. 2 A ). . Соответственно, уровни транскриптов маркерных генов AM LjHA1 , LjPT4 и RAM2 были значительно снижены у обоих мутантов по сравнению с диким типом (фиг. 2 B ). Несмотря на сильно сниженные уровни транскрипта CBX1 , экспрессия маркерного гена все еще индуцируется у мутантов (рис.2 B ), что предполагает наличие функционально дублирующих регуляторов. Применение фосфатов подавляет микоризацию и индуцированные микоризой факторы транскрипции (34, 38, 39). Сниженная экспрессия маркерного гена у мутанта cbx1-2 была более выражена при сдвиге Pi (500 мкМ) ( SI Приложение , фиг. S4 C и D ). Кроме того, сверхэкспрессия CBX1 привела к повышенному уровню микоризации и экспрессии LjPT4 ( SI Приложение , рис.S4 E ). В итоге результаты предполагают, что CBX1 участвует в образовании арбускул и экспрессии генов-хозяев в функционировании PAM.
CBX1 необходим для микоризной колонизации. ( A ) Скорость микоризации в мутантных линиях Gifu-129, cbx1-2 и cbx1-3 , выращенных при низком Pi (100 мкМ) в присутствии R. irregularis . А, арбускулы; H, гифы; V — пузырьки; A + H (%), процент корневых секторов с арбускулами и гифами; A + V + H (%), процент корневых секторов с арбускулами, пузырьками и гифами; H (%) — процент корневых секторов только с гифами; V + H (%), процент корневых секторов с пузырьками и гифами.Показано среднее значение ± стандартное отклонение ( n = 3). Использовался однофакторный дисперсионный анализ с последующим тестом достоверно значимой разницы Тьюки (HSD) [ F (A + H) 2,6 = 9,261; F (V + H) 2,6 = 9,874; F (A + V + H) 2,6 = 70,54; F (Всего) 2,6 = 73,2; P <0,05]. Разные буквы обозначают разные статистические группы. n.s., не имеет значения. ( B ) Экспрессия гена микориз в cbx1 аллельных мутантах в отсутствие (-) или в присутствии (+) R.irregularis ( n = 3). Односторонний дисперсионный анализ с последующим тестом HSD Тьюки [ F ( CBX1 ) 5,12 = 30,73; F ( LjHA1 ) 5,12 = 37,54; P <0,05] и непараметрический эквивалент критерия Краскела – Уоллиса [χ 2 ( RAM2 ) = 15,082; χ 2 ( LjPT4 ) = 14,342; df = 5, P <0,05] были использованы для определения значимости. Этот эксперимент был независимо повторен трижды с аналогичными результатами.( C и E ) pCBX1: активация GUS в корнях L. japonicus в присутствии R. irregularis . ( D и F ) Окрашивание WGA488 грибковых структур AM в тех же секторах. (Масштабные линейки: C и D, 500 мкм; E и F , 200 мкм.)
Разные буквы обозначают разные статистические группы. n.s., не имеет значения. ( B ) Экспрессия гена микориз в cbx1 аллельных мутантах в отсутствие (-) или в присутствии (+) R.irregularis ( n = 3). Односторонний дисперсионный анализ с последующим тестом HSD Тьюки [ F ( CBX1 ) 5,12 = 30,73; F ( LjHA1 ) 5,12 = 37,54; P <0,05] и непараметрический эквивалент критерия Краскела – Уоллиса [χ 2 ( RAM2 ) = 15,082; χ 2 ( LjPT4 ) = 14,342; df = 5, P <0,05] были использованы для определения значимости. Этот эксперимент был независимо повторен трижды с аналогичными результатами.( C и E ) pCBX1: активация GUS в корнях L. japonicus в присутствии R. irregularis . ( D и F ) Окрашивание WGA488 грибковых структур AM в тех же секторах. (Масштабные линейки: C и D, 500 мкм; E и F , 200 мкм.)
CBX1 трансактивирует
LjPT4 в зависимости от CTTC CRE. Чтобы проверить транскрипционную активность CBX1, мы изучили его субклеточную локализацию и активность промотора CBX1 .Слитые белки CBX1-YFP и GFP-CBX1 локализованы исключительно в ядре трансгенных волосатых корней культивируемых клеток L. japonicus и Arabidopsis ( SI Приложение , рис. S5 A и C ). Гистохимический анализ GUS CBX1 размером 1,9 т.п.н. в трансгенных волосатых корнях показал клеточно-автономную экспрессию CBX1 исключительно в секторах корня, колонизированных грибком AM R. irregularis (рис.2 C — F ). ).Более того, паттерны экспрессии генов CBX1 и LjPT4 коррелировали в разных типах органов растений с различным микоризным статусом ( SI, приложение , рис. S5 B ). Затем слитый белок CBX1, меченный GFP, был коэкспрессирован с pLjPT4: GUS или pLjPT4 — mCTTC : репортерами GUS или с репортерами 4 * CTTC: GUS или GUS : , соответственно, в культивируемых в суспензии корневых клетках микоризного nonhost A. thaliana (рис.3 A ). Повышенное накопление красителя индиго (продукта активности GUS) в клетках указывало на активацию промотора LjPT4 с помощью CBX1, в то время как промотор LjPT4 , содержащий mCTTC, не был активирован (фиг. 3 B ). Слияние GFP-CBX1 трансактивировало химерный ген 4 * CTTC: GUS , но не его мутированную версию ( 4 * mCTTC: GUS ), в то время как GFP сам по себе не влиял на репортерную систему.
thaliana (рис.3 A ). Повышенное накопление красителя индиго (продукта активности GUS) в клетках указывало на активацию промотора LjPT4 с помощью CBX1, в то время как промотор LjPT4 , содержащий mCTTC, не был активирован (фиг. 3 B ). Слияние GFP-CBX1 трансактивировало химерный ген 4 * CTTC: GUS , но не его мутированную версию ( 4 * mCTTC: GUS ), в то время как GFP сам по себе не влиял на репортерную систему.
CBX1 регулирует гены микоризных маркеров у разных видов двудольных.( A ) Схема репортера и эффектора, используемых в анализе трансактивации. DsRed использовался для проверки эффективности преобразования. p35S: GFP , отрицательный контроль. ( B ) Анализ трансактивации с факторами транскрипции AP2 на четырех химерных репортерных генах. Окрашивание GUS культивированных в суспензии клеток показано в верхней части графика. Среднее ± стандартное отклонение ( n = 3). Был проведен односторонний дисперсионный анализ с последующим HSD Тьюки ( F 27,56 = 44,38, P <0.001). ( C ) Сверхэкспрессия CBX1 увеличивала экспрессию генов LjPT4 , LjHA1 и RAM2 в L. japonicus в отсутствие AM-грибов. Пределы прямоугольника указывают на 25-й и 75-й процентили. Усы столбикового графика простираются до значения, которое не превышает 1,5-кратного интерквартильного размаха от верхнего или нижнего квартиля. Выбросы нанесены точками. Использовался тест Стьюдента t ( n = 7). ( D ) Эктопическая экспрессия CBX1 в волосатых корнях картофеля приводила к накоплению транскриптов индуцированных микоризой переносчиков Pi и генов H + -АТФазы.Использовался тест Стьюдента t ( n = 3). ( E ) Индукция генов, связанных с MUP, при эктопической экспрессии CBX1 в листьях табака. Использовался тест Стьюдента t ( n = 6). * P <0,05; ** P <0,01; *** P <0,001. Было проведено три независимых эксперимента с аналогичными результатами. ( F ) Трансактивация CBX1 генов переносчиков Pi из разных видов растений и L. japonicus LjHA1 и RAM2 в A.thaliana суспензия культивируемых клеток. Показаны средние значения ± SD активности GUS из трех биологических повторов ( n = 3; тест Стьюдента t ; * P <0,05; *** P <0,001). Этот эксперимент был повторен трижды независимо с аналогичными результатами.
Было проведено три независимых эксперимента с аналогичными результатами. ( F ) Трансактивация CBX1 генов переносчиков Pi из разных видов растений и L. japonicus LjHA1 и RAM2 в A.thaliana суспензия культивируемых клеток. Показаны средние значения ± SD активности GUS из трех биологических повторов ( n = 3; тест Стьюдента t ; * P <0,05; *** P <0,001). Этот эксперимент был повторен трижды независимо с аналогичными результатами.
Чтобы показать, что активация промотора специфична для CBX1, три фактора транскрипции AP2, WRI5a, WRI5c и WRI3, слитые с GFP, также коэкспрессируются с репортерными конструкциями GUS в культивируемых в суспензии клетках.За исключением CBX1, все три белка AP2 не смогли активировать экспрессию pLjPT4: GUS или 4 * CTTC: GUS (фиг. 3 B ). Мы также обнаружили, что два карбоксиконцевых усечения CBX1 1–221 и CBX1 1–308 , которые сохранили способность связываться с мотивом CTTC in vitro (рис. 1 F ) и локализовались в ядре ( SI Приложение , рис. S5 C ), не удалось активировать LjPT4 и синтетический промотор 4 * CTTC в A.thaliana (рис.3 B ). Эти результаты показали, что трансактивация промотора LjPT4 зависела от присутствия карбоксиконцевой кислотной области CBX1, потенциального домена трансактивации, включающего аминокислоты 309–378 (40).
CBX1-CTTC Механизм регуляции CRE сохраняется у видов-хозяев AM.
Основываясь на предложенной модульной конструкции AM-симбиоза (41), мы предположили, что CBX1 регулирует генный модуль для контроля микоризного транспорта питательных веществ.Чтобы проверить эту гипотезу корегуляции, CBX1 ( pUB: CBX1-YFP ) эктопически экспрессировался в волосатых корнях трансгенных L. japonicus в отсутствие AM-грибов, что привело к значительному увеличению уровня LjPT4 транскриптов относительно контроля, тогда как LjPT1 , LjPT2 и LjPT3 остались неизменными (фиг. 3 C ). Протон-АТФаза (НА1), локализованная в мембране, важна для MPU за счет активации протон-связанного транспорта Pht1 Pi (18, 19, 42).Экспрессия LjHA1 была индуцируемой микоризой в корнях ( SI, приложение , фиг. S6, A ), и ее промоторная область, содержащая мотив CTTC, была напрямую связана с CBX1 ( SI, приложение , фиг. S6 B ). Соответственно, сверхэкспрессия CBX1 в трансформированных корнях приводила к значительному накоплению транскриптов LjHA1 в отсутствие AM-грибов. Кроме того, эктопическая экспрессия L. japonicus CBX1 в трансгенных волосатых корнях Solanum tuberosum и в листьях Nicotiana benthamiana привела к значительному накоплению мРНК, кодирующей соответствующий индуцируемый микоризой переносчик PiATPase и H + , соответственно (рис.3 D и E ). В общем, это указывает на сохранение цис- -регулирующей активности CBX1 в поглощении микоризным Pi у различных видов эвдикотов.
3 C ). Протон-АТФаза (НА1), локализованная в мембране, важна для MPU за счет активации протон-связанного транспорта Pht1 Pi (18, 19, 42).Экспрессия LjHA1 была индуцируемой микоризой в корнях ( SI, приложение , фиг. S6, A ), и ее промоторная область, содержащая мотив CTTC, была напрямую связана с CBX1 ( SI, приложение , фиг. S6 B ). Соответственно, сверхэкспрессия CBX1 в трансформированных корнях приводила к значительному накоплению транскриптов LjHA1 в отсутствие AM-грибов. Кроме того, эктопическая экспрессия L. japonicus CBX1 в трансгенных волосатых корнях Solanum tuberosum и в листьях Nicotiana benthamiana привела к значительному накоплению мРНК, кодирующей соответствующий индуцируемый микоризой переносчик PiATPase и H + , соответственно (рис.3 D и E ). В общем, это указывает на сохранение цис- -регулирующей активности CBX1 в поглощении микоризным Pi у различных видов эвдикотов.
Близкие гомологи CBX1 существуют в разных таксонах ( SI Приложение , Рис. S7). Для проверки гипотезы о том, что механизм регуляции транскрипции, контролируемый CBX1, является эволюционно консервативным, промоторы из генов Pht1, индуцируемых микоризой, LjPT3 , LjPT4 , MtPT4 , картофеля StPT3 4 и 1962 StPT3 4 и 62 StPar PtPT12 , OsPT11 и ZmPT6 были слиты с репортерным геном GUS и котрансформированы с CBX1 в A.thaliana суспензионно культивируемые клетки корней (4, 9, 11, 12, 33, 43⇓ – 45). За исключением OsPT11 , все остальные восемь генов содержат в своих промоторах мотивы CTTC. В этой системе in vivo CBX1 активировал промоторы LjPT4 , StPT4 , MtPT4 и PtPT10 из генов подсемейства I Pht1, но не промоторы из однодольных, таких как2 OsPT11 или субсемейства Pht11 и . III, включая StPT3 , LjPT3 и PtPT12 (рис.3 F ). Эта активация специфических промоторов из микоризных хозяев эвдикота объясняет предыдущие результаты, полученные с промотором OsPT11 из риса, который не активировался при трансформации в микоризный картофель и корни M. truncatula (5). CBX1 также индуцировал экспрессию GUS, управляемую промотором гена LjHA1 (фиг. 3 F ). В целом эти данные предполагают действие микоризного генного модуля, включающего CBX1, LjPT4 и LjHA1 , участвующих в MPU у растений эвдикота.
III, включая StPT3 , LjPT3 и PtPT12 (рис.3 F ). Эта активация специфических промоторов из микоризных хозяев эвдикота объясняет предыдущие результаты, полученные с промотором OsPT11 из риса, который не активировался при трансформации в микоризный картофель и корни M. truncatula (5). CBX1 также индуцировал экспрессию GUS, управляемую промотором гена LjHA1 (фиг. 3 F ). В целом эти данные предполагают действие микоризного генного модуля, включающего CBX1, LjPT4 и LjHA1 , участвующих в MPU у растений эвдикота.
Общегеномные мишени CBX1.
В дополнение к LjPT4 и LjHA1 , количество транскриптов RAM2 , кодирующее глицерин-3-фосфатацилтрансферазу, необходимое для развития арбускул (46), было значительно увеличено (фиг.3 C ), в то время как экспрессия других микоризных гены-маркеры, такие как SbtM1 , STR и BCP1 , не были затронуты (5, 47⇓ – 49). CBX1 также активировал промотор гена RAM2 (рис.3 F ), предполагая, что CBX1 управляет экспрессией широкого спектра генов, участвующих в развитии AM-симбиоза. Поэтому мы исследовали глобальные сайты связывания ДНК слитого белка CBX1-YFP в геноме L. japonicus с использованием иммунопреципитации хроматина в сочетании с высокопроизводительным секвенированием (ChIP-seq) ( SI Приложение , рис. S8 A и В ). Всего 136 генов-мишеней принадлежали к общему пересечению в двух повторностях, что указывает на высокую значимость перекрытия между повторностями (точный критерий Фишера, отношение шансов 184.95, P <2.2 e -16; Набор данных S1). Сайты связывания CBX1 были обогащены рядом с сайтом начала транскрипции генов-мишеней (фиг. 4 A ) и преобладали в промоторах, 5′-UTR и межгенных областях (фиг. 4 B ). Анализ функциональной аннотации показал, что гены, участвующие в метаболизме и транскрипции липидов, составляют большую часть из 136 мишеней CBX1, помимо небелковых кодирующих и неизвестных генов ( SI Приложение , рис. S8 C ). Интеграция нашего анализа ChIP-seq со сравнительным анализом L.japonicus и Регуляция гена R. irregularis при разрешении транскриптов (RNA-seq) (Dataset S2) (50) привела к 43 мишеням CBX1, которые совпадали с индуцируемыми микоризой генами (точный критерий Фишера, отношение шансов 17,53, P <2.2 e -16) (рис.4 C и набор данных S1), включая LjPT4 (рис.4 D ). ChIP-qPCR подтвердила, что слитый белок CBX1-YFP обладал способностью осаждать область промотора LjPT4 , содержащую CTTC CRE (рис.4 E и F ). CTTC-содержащий область в LjPT3 не была обогащена ни CFP, ни CBX1-YFP, что подтвердило результат ChIP-seq. В соответствии с гомологом WRI1 A. thaliana (22, 23), 12 из 43 генов-мишеней CBX1, индуцируемых микоризой, относятся к метаболизму липидов и включают девять генов, участвующих в биосинтезе жирных кислот [субъединица пируватдегидрогеназы E1 ( PDH_E1 α_ и β), дигидролипоилдегидрогеназа 1 ( LPD1 ), белок-носитель карбоксила биотина 2 ( BCCP2 ), α-карбоксилтрансфераза (α -CT ), трансацилаза малонил-CoA-ACP ( MAT 1), ацильный белок-носитель ( ACP1 ), еноил-ACP-редуктаза ( ENR ) и ацил-ACP-тиоэстераза ( FatM )] и три гликолитических гена [глицерин-3-фосфатдегидрогеназа ( GPDH трансфокатор ), фосфоенолпирув62 PPT ) и изофермент G пируваткиназы ( PK )] (рис.4 G и SI Приложение , рис. S9). Недавние исследования подчеркнули важную роль синтеза жирных кислот 16: 0 в микоризных растениях-хозяевах и его предполагаемую передачу AM-грибам для поддержания симбиоза (26-29, 51). Микоризная индукция 11 липид-связанных генов в M. truncatula зависит от активности регулятора GRAS RAM1 (29). Восемь из этих генов были мишенями CBX1 в L. japonicus , включая BCCP2 , PDH_E1 β, PK , ACP1 , MAT , ENR , GPDH и . RAM2 и LjHA1 не были включены в наш список 136 мишеней из-за неполной последовательности генома или присутствия только в одном повторном эксперименте ( SI Приложение , рис. S13 C ). Поэтому мы вручную добавили последовательность RAM2 к последовательности генома L. japonicus , и считывания короткой последовательности ChIP-seq были достаточны для точного картирования обогащенных фрагментов ДНК на RAM2 ( SI Приложение , рис. S9) . В итоге результаты показали, что CBX1 обладает способностью регулировать гены, лежащие в основе различных функций AM, включая MPU и биосинтез жирных кислот.
S8 C ). Интеграция нашего анализа ChIP-seq со сравнительным анализом L.japonicus и Регуляция гена R. irregularis при разрешении транскриптов (RNA-seq) (Dataset S2) (50) привела к 43 мишеням CBX1, которые совпадали с индуцируемыми микоризой генами (точный критерий Фишера, отношение шансов 17,53, P <2.2 e -16) (рис.4 C и набор данных S1), включая LjPT4 (рис.4 D ). ChIP-qPCR подтвердила, что слитый белок CBX1-YFP обладал способностью осаждать область промотора LjPT4 , содержащую CTTC CRE (рис.4 E и F ). CTTC-содержащий область в LjPT3 не была обогащена ни CFP, ни CBX1-YFP, что подтвердило результат ChIP-seq. В соответствии с гомологом WRI1 A. thaliana (22, 23), 12 из 43 генов-мишеней CBX1, индуцируемых микоризой, относятся к метаболизму липидов и включают девять генов, участвующих в биосинтезе жирных кислот [субъединица пируватдегидрогеназы E1 ( PDH_E1 α_ и β), дигидролипоилдегидрогеназа 1 ( LPD1 ), белок-носитель карбоксила биотина 2 ( BCCP2 ), α-карбоксилтрансфераза (α -CT ), трансацилаза малонил-CoA-ACP ( MAT 1), ацильный белок-носитель ( ACP1 ), еноил-ACP-редуктаза ( ENR ) и ацил-ACP-тиоэстераза ( FatM )] и три гликолитических гена [глицерин-3-фосфатдегидрогеназа ( GPDH трансфокатор ), фосфоенолпирув62 PPT ) и изофермент G пируваткиназы ( PK )] (рис.4 G и SI Приложение , рис. S9). Недавние исследования подчеркнули важную роль синтеза жирных кислот 16: 0 в микоризных растениях-хозяевах и его предполагаемую передачу AM-грибам для поддержания симбиоза (26-29, 51). Микоризная индукция 11 липид-связанных генов в M. truncatula зависит от активности регулятора GRAS RAM1 (29). Восемь из этих генов были мишенями CBX1 в L. japonicus , включая BCCP2 , PDH_E1 β, PK , ACP1 , MAT , ENR , GPDH и . RAM2 и LjHA1 не были включены в наш список 136 мишеней из-за неполной последовательности генома или присутствия только в одном повторном эксперименте ( SI Приложение , рис. S13 C ). Поэтому мы вручную добавили последовательность RAM2 к последовательности генома L. japonicus , и считывания короткой последовательности ChIP-seq были достаточны для точного картирования обогащенных фрагментов ДНК на RAM2 ( SI Приложение , рис. S9) . В итоге результаты показали, что CBX1 обладает способностью регулировать гены, лежащие в основе различных функций AM, включая MPU и биосинтез жирных кислот.
Полногеномная идентификация генов-мишеней CBX1 с помощью ChIP-seq. ( A ) Анализ ChIP-seq показывает пики связывания CBX1, обогащенные рядом с сайтом начала транскрипции (TSS). Использовали пики, общие для двух повторных экспериментов. Иммунопреципитированные фрагменты ДНК из волосистых корней возрастом 1 месяц, несущие pUB: CBX1-YFP или pUB: CFP отрицательный контроль, подвергали секвенированию ДНК. ( B ) Распределение 136 сайтов связывания CBX1 в геноме L. japonicus .( C ) Диаграмма Венна, изображающая перекрытие между мишенями CBX1 из ChIP-seq и генов, регулируемых микоризой. Уникальные гены (392 и 226) были значительно обогащены CBX1 в двух экспериментах (в два раза выше в окружающей области размером 10 т.п.н.; кратное изменение относительно контроля CFP> 2; P <0,0001). повторить, повторить. ( D ) Просмотр в браузере IGV связывания CBX1 с геном LjPT4 . Дорожки отображают данные из образцов Input, ChIPed CFP и ChIPed CBX1 (две реплики).Число в верхнем левом углу каждой дорожки указывает высоту дорожки (300 показаний на ячейку). Пик, идентифицированный у Гомера, обозначен синей полосой. Толстые линии представляют экзоны, а тонкие линии интроны в структуре гена. Черная стрелка указывает TSS. Синие и красные галочки под структурой гена указывают на ядерную последовательность CTTC (TCTTGT) или AW-бокс (CnTnG (n) 7 CG) на положительной и отрицательной цепи ДНК соответственно. ( E ) Схематическое изображение областей генома LjPT4 и LjPT3 в масштабе.Черные полосы представляют кодовую область. Линии представляют собой некодирующую ДНК. Блок CTTC CRE и AW обозначены в промоторных областях черными или красными полосами соответственно. P1-P4 представляют собой фрагменты ДНК, предназначенные для ChIP-qPCR. ( F ) Подтверждение ChIP-seq с помощью ChIP-qPCR, что CBX1 связывается с промотором LjPT4 . После нормализации с вводом вычисляли кратное обогащение по сравнению с CFP, содержащим чипы анти-GFP. Показаны средние значения ± стандартное отклонение трех независимых биологических повторов. Использовался тест Стьюдента t .* P <0,05. ( G ) Гены, индуцируемые микоризой, связанные с липидами, были нацелены CBX1 в ChIP-seq. Тепловая карта профилей экспрессии гена-мишени CBX1 на основе log 10 преобразованных импульсов на миллион (имп / мин), отображенных на основе анализа данных RNA-seq (50). Количество бокса AW и ядра CTTC (TCTTGT) подсчитывали в пиках гомера из ChIP-seq. ( H ) Накопление транскриптов CBX1-мишеней в волосатых корнях L. japonicus , сверхэкспрессирующих CBX1, в отсутствие AM-грибов.Использовался тест Стьюдента t ( n = 7). * P <0,05; ** P <0,01; *** P <0,001. Три эксперимента были выполнены независимо с аналогичными результатами. ( I ) Анализ трансактивации с CBX1 на pLjPT4: репортер GUS с мутациями на CTTC или / и AW боксе. Среднее ± стандартное отклонение ( n = 3). Был проведен односторонний дисперсионный анализ с последующим HSD Тьюки ( F 15,32 = 14,17, P <0,05).
AW-бокс обогащен CBX1-связанными сайтами липидных метаболических генов (фиг. 4 G, и SI, приложение , фиг. S9). CBX1 непосредственно связывается с боксом AW in vitro ( SI, приложение , фиг. S10, A ), что указывает на консервативные связывающие свойства гомологов WRI. Сверхэкспрессия CBX1 значительно увеличивала уровни транскриптов BCCP2 , PDHC_E1a , PDHC_E1 β, LPD1 , ENR , GPDH , ABCB , K1962 и Fat japonicus (рис.4 H ). Аналогичным образом, повышенное количество транскриптов генов биосинтеза жирных кислот также наблюдалось при эктопической экспрессии CBX1 в листьях табака и в волосатых корнях картофеля ( SI Приложение , рис. S10 B и C ), которые стояли в соответствует специфическому накоплению триацилглицеринов в листьях табака после эктопической экспрессии L. japonicus CBX1, хотя общее содержание жирных кислот не изменилось ( SI Приложение , рис.S11 A и B ). В CBX1 оба домена AP2 и область 212–308 необходимы для связывания с AW box и CTTC (Fig. 1 E и F и SI Приложение , Fig. S12 A ). Более того, AW-бокс действует во взаимодействии с CTTC в связывании промотора LjPT4 in vitro и его трансактивации in vivo с помощью CBX1 (фиг. 4 I и SI, приложение , фиг. S12 B ). В итоге результаты предполагают, что CBX1-направленная AM-специфическая регуляция гена посредством прямого связывания с CTTC и AW box в регуляторной области различных генов-мишеней.
Помимо MPU и липидных генов, транскрипты трех генов GRAS, кодирующих гомологи M. truncatula MIG1 (Lj6g3v1
0.1) (52) и MIG1-подобных белков (Lj5g3v1598410.1 и Lj1g3v4851630.1), были обогащены на CBX1 ( SI Приложение , рис. S13 A и B ), который поместил CBX1 в регуляторную сеть генов AM-симбиоза. В целом, наши результаты показали, что CBX1 корегулирует различные генные модули благодаря своей способности распознавать два мотива дивергентных последовательностей, которые опосредуют такие функции, как MPU и биосинтез жирных кислот, а также другие, еще плохо изученные взаимосвязанные процессы, участвующие в симбиозе AM.Обсуждение
Взаимный обмен питательными веществами стабилизирует сотрудничество между микобионтом и фитобионтом в симбиозе AM в течение эволюционного времени (53). Что касается теории «биологического рынка» (53, 54), регуляторы, участвующие в оркестровке биологических процессов, лежащих в основе мутуализма, вероятно, играли важную роль в эволюции AM-симбиоза. Мы показываем здесь, что индуцируемый микоризой CBX1 из L. japonicus , фактор транскрипции WRI1, действует как регулятор и активирует гены, кодирующие специфичный для микоризы переносчик Pi и протон-АТФазу из различных растений эвдикота и белков, участвующих в биосинтезе жирных кислот.Вычислительная идентификация консервативного CTTC CRE (TCTTGTT) (рис.1 C ) (14) посредством межвидового сравнения генов, регулируемых микоризой, соответствовала специфичности связывания CBX1 с основной последовательностью TCTTGT, показанной с помощью EMSA (рис. 1 D ). Анализы трансактивации в суспензионных культивируемых клетках немикоризного хозяина Исследования избыточной экспрессии A. thaliana и CBX1 в трансформированных корнях или листьях L. japonicus , картофеля и табака в отсутствие AM-грибов предполагают наличие консервативного регуляторного механизма, контролирующего одновременное экспрессия генов подсемейства I Pht1 и генов протон-АТФазы у эвдикотов.Неспособность CBX1 связываться in vivo (фиг.4 F ) и не активировать (фиг.3 F ) промотор LjPT3 , который также содержит мотив CTTC, предполагает важную роль последовательностей, фланкирующих мотив CTTC в cis регламент. Значительно сниженная, но не устраненная активность GUS, управляемая LjPT4 -mCTTC в трансгенных корнях, также указывает на существование альтернативных CRE (фиг. 1 A ). Взаимодействие мотива CTTC и AW-бокса было обнаружено в промоторах нескольких микориз-специфичных и активируемых микоризой генов (рис.4 G ). CBX1 мог связываться с обоими мотивами (фиг.1 и SI Приложение , фиг. S9, S10 A и S12), и CBX1-опосредованная активация LjPT4 зависела от обоих мотивов (фиг.4 I ), подразумевая, что два мотива строят молекулярный модуль в генах симбиоза AM. Точная регуляторная функция CBX1 на молекулярном модуле CTTC / AW индивидуального гена-мишени требует дальнейшего изучения. Сохраняющаяся экспрессия гена микоризы у мутантов cbx1 (рис.2 B ) предположили, что другие факторы транскрипции могут функционировать избыточно, такие как AM-индуцибельный WRI5a / b / c (29, 34) с CTTC-связывающей способностью ( SI, приложение , рис. S2).
Полногеномная идентификация сайтов связывания CBX1 с помощью ChIP-seq выявила 43 индуцируемых микоризой мишени CBX1 (набор данных S1). Среди них 12 генов участвуют в синтезе жирных кислот и гликолизе de novo. Обогащение AW-бокса этими связывающими областями поддерживает консервативную регуляцию липидного синтеза с помощью WRI-подобных белков у различных видов растений (23, 55).В списке из 43 генов гены, кодирующие LjPT4, FatM (AMP-зависимую синтетазу и лигазу) и белок семейства ABC-транспортеров B ( ABCB ), были консервативны у филогенетически различных видов микоризных хозяев (56). Кроме того, мы показали, что LjHA1 напрямую регулируется CBX1, поскольку CBX1 обладал способностью связывать TCTTGT-содержащую промоторную область LjHA1 in vitro ( SI Приложение , рис. S6 B ) и активировал pLjHA1: химерный ген GUS в A.thaliana (рис.3 F ). Две цели CBX1 RAM2 и LjHA1 изначально не были идентифицированы в нашем анализе ChIP-seq, который предполагает, что некоторые цели были пропущены из-за неполной информации о геноме или ошибок аннотации, технических препятствий или жестких критериев для выбора пиков через Трубопровод Гомера ( SI Приложение , рис. S9 и S13 C ). Кроме того, MIG1 , регулятор экспансии кортикальных клеток, необходимый для развития арбускул (52), был идентифицирован как ген-мишень CBX1 с помощью ChIP-seq и ChIP-qPCR ( SI Приложение , рис.S13 A и B ). В M. truncatula усеченный промотор длиной 230 пар оснований MIG1 , содержащий два мотива CTTC, но не содержащий AW-бокса, был достаточен для управления экспрессией GUS в ответ на AM-грибы (52). Поскольку сверхэкспрессия CBX1 использовалась для ChIP-seq в отсутствие грибов AM, дальнейшие исследования подтвердят функцию конкретных мишеней CBX1 в микоризном симбиозе. Было показано, что гены
LjPT4 , LjHA1 и RAM2 регулируются белком GRAS RAM1 (29, 57, 58), который непосредственно регулируется комплексом CCaMK – CYCLOPS – DELLA (49).Требуются дополнительные исследования, чтобы выяснить, участвует ли CBX1 в экспрессии микоризных генов независимо, совместно или ниже RAM1 во время развития AM. Продолжение исследований того, как CBX1 и ортологичные белки произошли от ранних наземных растений и их водорослевых предков (41), помогут понять эволюцию регуляторных модулей, которые определяют мутуалистические взаимодействия на границе корень-гриб в симбиозе AM.
Материалы и методы
Подробная информация о растительных материалах и условиях роста представлена в Приложении SI .Трансформация волосистых корней и листьев, очистка белка и EMSA, количественный анализ ПЦР в реальном времени, филогенетический анализ, гистохимический анализ GUS, анализ трансактивации, локализация субклеточного белка, ChIP-seq и анализ данных RNA-seq описаны в приложении SI . , С.И. Материалы и методы . Конструкции и праймеры перечислены в приложении SI , таблицы S1 – S6. Цели CBX1 и их функциональная аннотация перечислены в наборе данных S1. Гены, реагирующие на R.irregularis в L. japonicus были идентифицированы с использованием RNA-seq, как описано ранее (50), и перечислены в Dataset S2.
Благодарности
Мы благодарим доктора К. Шлюкинга и В. Вевер, Ю. Арльта и К. Нотелле за экспериментальную поддержку; Доктору И. Фабианской за помощь в статистическом анализе данных; Доктору М. Бёмеру (Университет Мюнстера) за поддержку EMSA; Д-ру Н. Герлаху за предоставление плазмиды pRedRoot-pZmPT6: GUS ; Д-ру Ф. Мартину (Национальный институт агрономических исследований) за предоставление геномной ДНК тополя; ДоктораФ. Хе и У. Хеккера за полезные обсуждения; и С. Верт за фотографии. Это исследование было поддержано исследовательской стипендией фонда Александра фон Гумбольдта (LX), грантом Института содействия преподаванию науки и технологий Таиланда (LK), Международной исследовательской школой Макса Планка на тему «Понимание сложных свойств растений с использованием вычислительных и Эволюционные подходы »в Кельне (GS) и Грант Немецкого научного фонда BU-2250 / 12-1 (М. Бухеру)
Footnotes
Авторские вклады: L.X. и M. Bucher разработали исследование; L.X., L.K., Y.Z., V.V. и M. Brands проводили исследования; T.G. внесены новые реагенты / аналитические инструменты; L.X., L.K., G.S., M. Brands, P.D., F.T. и M. Bucher проанализировали данные; и L.X. и М. Бухер написал статью.
Авторы заявляют об отсутствии конфликта интересов.
Эта статья представляет собой прямое представление PNAS.
Эта статья содержит вспомогательную информацию на сайте www.pnas.org/lookup/suppl/doi:10.1073 / pnas.1812275115 / — / DCSupplemental.
- Copyright © 2018 Автор (ы). Опубликовано PNAS.
(PDF) Обмен гистонов, структура хроматина и регуляция транскрипции
41. Petruk, S. etal. Белки TrxG и PcG, но не метилированные гистоны
, остаются связанными с ДНК
посредством репликации. Cell 150, 922–933 (2012).
42. Williams, S.K., Truong, D. И Тайлер, J.K. Ацетилирование в глобулярном ядре гистона h4
на лизине 56 способствует разборке хроматина
во время транскрипционной активации
.Proc. Natl Acad. Sci. USA 105,
9000–9005 (2008).
43. Тропбергер, P. etal. Регуляция транскрипции
посредством ацетилирования h4K122 на боковой поверхности
октамера гистонов. Cell 152, 859–872 (2013).
44. Шогрен-Кнаак, M. etal. Ацетилирование гистона h5K16
контролирует структуру хроматина и белковые взаимодействия.
Наука 311, 844–847 (2006).
45. Lee, J.S. etal. Пересечение гистонов между моноубиквитинированием h3B
и метилированием h4, опосредованным
COMPASS.Cell 131, 1084–1096 (2007).
46. Венкатеш, S. etal. Set2 метилирование гистона h4
лизин 36 подавляет обмен гистонов на транскрибированных генах
. Nature 489, 452–455 (2012).
47. Папамихос-Хронакис, M., Ватанабе, S., Rando, O.J.
& Peterson, C.L. Глобальная регуляция локализации h3A.Z
с помощью фермента INO80, ремоделирующего хроматин
, важна для целостности генома. Cell 144,
200–213 (2011).
Эта статья характеризует роль комплекса INO80
в предотвращении неправильной локализации варианта
h3A.Z в областях генома, отличных от
промотора.
48. McKittrick, E., Gafken, P.R., Ahmad, K. И Хеникофф, S.
Гистон h4.3 обогащен ковалентными модификациями
, связанными с активным хроматином. Proc. Natl Acad. Sci.
USA 101, 1525–1530 (2004).
49. Тропбергер, P. И Шнайдер, R.Почесывание (латеральной) поверхности регуляции хроматина
модификациями гистона
. Структура природы. Мол. Биол. 20, 657–661
(2013).
50. Тессарз, П. И Кузаридес, T. Модификации гистонового ядра
, регулирующие структуру и динамику нуклеосом.
Nature Rev. Mol. Cell Biol. 15. С. 703–708 (2014).
51. Becker, P.B. И Уоркман, J.L. Ремоделирование и эпигенетика нуклеосомы
. Харб Холодного источника.
Перспектива.Биол. 5, а017905 (2013).
52. Кобор, M.S. etal. Белковый комплекс, содержащий
консервативную Swi2 / Snf2-родственную АТФазу Swr1p, откладывает
варианта гистона h3A.Z в эухроматин. PLoS Biol. 2,
E131 (2004).
53. Watanabe, S. И Петерсон, C.L. Семейство INO80 ферментов ремоделирования хроматина
: регуляторы динамики вариантов гистона
. Харб Холодного источника. Symp. Quant.
Биол. 75, 35–42 (2010).
54.Мидзугути, G. etal. АТФ-управляемый обмен варианта h3AZ гистона
, катализируемый ремоделирующим комплексом SWR1 хроматина
. Science 303, 343–348
(2004).
55. Лук, E. etal. Пошаговая замена гистона на SWR1
требует двойной активации гистоном h3A.Z и канонической нуклеосомой
. Cell 143, 725–736
(2010).
56. Wu, W.H. etal. Swc2 представляет собой широко консервативный связывающий модуль h3AZ-
, необходимый для АТФ-зависимого обмена гистона
.Структура природы. Мол. Биол. 12, 1064–1071
(2005).
57. Watanabe, S., Radman-Livaja, M., Rando, O.J. &
Peterson, C.L. Переключатель ацетилирования гистонов регулирует отложение
h3A.Z с помощью фермента ремоделирования SWRC.
Science 340, 195–199 (2013).
58. Кущ, T. etal. Ацетилирование с помощью Tip60 необходимо для селективного обмена
вариантов гистона в повреждениях ДНК.
Science 306, 2084–2087 (2004).
59.Рул, D.D. etal. Очистка человеческого комплекса SRCAP
, который ремоделирует хроматин путем включения гистонового варианта
h3A.Z в нуклеосомы. Биохимия
45, 5671–5677 (2006).
60. Eissenberg, J.C., Wong, M. И Chrivia, J.C. Человеческий
SRCAP и Drosophila melanogaster DOM являются гомологами
, которые функционируют в сигнальном пути notch.
Мол. Клетка. Биол. 25, 6559–6569 (2005).
61. Cai, Y. etal.Белок YL1 млекопитающих является общей субъединицей
гистонацетилтрансферазы TRRAP / TIP60
и комплексов SRCAP. J.Biol. Chem. 280,
13665–13670 (2005).
62. Johnston, H., Kneer, J., Chackalaparampil, I.,
Yaciuk, P. & Chrivia, J. Идентификация нового члена семейства белков SNF2 /
SWI2, SRCAP, который взаимодействует
с CREB-связывающим белком. J.Biol. Chem. 274,
16370–16376 (1999).
63.Conaway, R.C. И Conaway, J.W. Комплекс ремоделирования хроматина INO80
в транскрипции,
репликации и репарации. Trends Biochem. Sci. 34,
71–77 (2009).
64. Yen, K., Vinayachandran, V. И Пью, B.F. SWRC и
ремоделирующие устройства хроматинаINO80 распознают свободные области
нуклеосом около +1 нуклеосомы. Cell 154,
1246–1256 (2013).
65. D’Arcy, S. etal. Шаперон Nap1 экранирует поверхности гистонов
, используемых в нуклеосомах, и может превращать h3A-h3B
в нетрадиционную тетрамерную форму.Мол. Cell 51,
662–677 (2013).
66. De Koning, L., Corpet, A., Haber, J.E. И Альмоузни, G.
Шапероны гистонов: сеть сопровождения, регулирующая трафик гистонов
. Структура природы. Мол. Биол. 14,
997–1007 (2007).
67. Hondele, M. etal. Структурная основа узнавания гистона h3A–
h3B эссенциальным шапероном FACT.
Природа 499, 111–114 (2013).
68. Bowman, A. etal. Гистоновые шапероны Nap1 и
Vps75 связывают гистоны h4 и h5 в тетрамерной конформации
.Мол. Cell 41, 398–408 (2011).
69. Swaminathan, V., Kishore, A.H., Febitha, K.K. &
Кунду, T.K. Шаперон гистонов человека нуклеофозмин
усиливает транскрипцию зависимого от ацетилирования хроматина
. Мол. Клетка. Биол. 25, 7534–7545 (2005).
70. Лук, E. etal. Chz1, ядерный шаперон для гистона
h3AZ. Мол. Cell 25, 357–368 (2007).
71. Вс, D. etal. Структурная основа распознавания h4K56-
ацетилированного гистона h4 – h5 шапероном Rtt106.
Nature 483, 104–107 (2012).
72. Owen-Hughes, T. И Уоркман, J.L. Ремоделирование структуры хроматина
,массива нуклеосом с помощью нацеленного на транскрипционный фактор
транс-смещениягистонов. EMBO J. 15, 4702–4712 (1996).
73. Lorch, Y., Maier-Davis, B. И Корнберг, R.D. Ремоделирование хроматина
путем разборки нуклеосом invitro.
Proc. Natl Acad. Sci. USA 103, 3090–3093 (2006).
74.Курян, Б.Г. etal. Плотность гистонов поддерживается
во время транскрипции, опосредованной ремоделером хроматина
RSC и гистоновым шапероном NAP1 in1vitro.
Proc. Natl Acad. Sci. USA 109, 1931–1936 (2012).
75. Han, J. etal. Rtt109 ацетилирует гистон h4 лизин 56
и участвует в репликации ДНК. Science 315,
653–655 (2007).
76. Driscoll, R., Hudson, A. И Джексон, S.P. Дрожжи Rtt109
способствуют стабильности генома за счет ацетилирования гистона h4
по лизину 56.Science 315, 649–652 (2007).
77. Rufiange, ,A., Jacques, P.E., Bhat, W., Robert, F. &
Нурани, A. Независимый от репликации по всему геному обмен
гистона h4 происходит преимущественно на промоторах
и вовлекает ацетилирование h4 K56 и
Asf1. Мол. Cell 27, 393–405 (2007).
78. Schwabish, M.A. И Struhl, K. Asf1 опосредует выселение и отложение гистона
во время элонгации с помощью РНК
полимеразы II.Мол. Cell 22, 415–422 (2006).
79. Колонько, E.M. etal. Каталитическая активация ацетилтрансферазы Rtt109 гистона
гистоновым шапероном.
Proc. Natl Acad. Sci. USA 107, 20275–20280
(2010).
80. Цубота, T. etal. Ацетилирование гистона h4K56
катализируется гистоновыми шаперон-зависимыми комплексами.
Мол. Cell 25, 703–712 (2007).
81. Каплан, T. etal. Регуляция включения h4K56ac в дрожжах, опосредованная клеточным циклом и шаперонами
.PLoS
Genet. 4, e1000270 (2008).
82. Youdell, M.L. etal. Роли Ctk1 и Spt6 в
, регулирующих различные состояния метилирования гистона
h4 лизина 36. Mol. Клетка. Биол. 28, 4915–4926
(2008).
83. Йох, S.M., Cho, H., Pickle, L., Evans, R.M. &
Jones, K.A. Домен Spt6 Sh3 связывает Ser2P
RNAPII для управления Iws1-зависимым сплайсингом мРНК и экспортом
. Genes Dev. 21. С. 160–174 (2007).
84. Du, H.N., Fingerman, I.M. И Бриггс, S.D. Метилирование гистона h4
K36 опосредуется путем метилирования трансгистона
, включающим взаимодействие между
Set2 и гистоном h5. Genes Dev. 22, 2786–2798
(2008).
85. Du, H.N. И Бриггс, S.D. Поверхность нуклеосомы, образованная остатками
гистона h5, h3A и h4, необходима для метилирования
собственного гистона h4 Lys36, ацетилирования гистона
и репрессии криптической транскрипции.
J.Biol. Chem. 285. С. 11704–11713 (2010).
86. Carrozza, M.J. etal. Метилирование гистона h4 с помощью Set2
направляет деацетилирование кодирующих областей с помощью Rpd3S, чтобы
подавляли ложную внутригенную транскрипцию. Cell 123,
581–592 (2005).
87. Kaplan, C.D., Laprade, L. И Уинстон, Ф. Транскрипция
факторов элонгации подавляют инициацию транскрипции из
криптических сайтов. Science 301, 1096–1099 (2003).
88.Хасан, A.H., Нили, K.E. И Уоркман, J.L.
Комплексы гистонацетилтрансферазы стабилизируют
Связывание SWI / SNF с промоторными нуклеосомами. Cell 104,
817–827 (2001).
89. Schneiderman, J.I., Orsi, G.A., Hughes, K.T.,
Loppin, B. & Ахмад, K. Истощенные нуклеосомами
хроматиновых промежутков привлекают факторы сборки для варианта гистона h4.3
. Proc. Natl Acad. Sci. USA 10 9,
19721–19726 (2012).
90.Дрю, H.R. И Трэверс, A.A. Изгиб ДНК и его отношение
к позиционированию нуклеосом. J.Mol. Биол. 186,
773–790 (1985).
91. Segal, E. etal. Геномный код для позиционирования нуклеосомы
. Nature 442, 772–778 (2006).
92. Hartley, P.D. И Мадхани, H.D. Механизмы
определяют расположение и идентичность промоторных нуклеосом.
Cell 137, 445–458 (2009).
93. Гангули, О.Д., Череджи, И.Р.В., Ибен, О.Дж.R., Cole, H.A. &
Clark, D.J. RSC-зависимая конструктивная и
деструктивная интерференция между противостоящими массивами
фазированных нуклеосом в дрожжах. Genome Res. 24,
1637–1649 (2014).
94. Ranjan, A. etal. Область, свободная от нуклеосом, доминирует над ацетилированием гистона
при нацеливании SWR1 на промоторы
для замены h3A.Z. Cell 154, 1232–1245
(2013).
Ссылки 54–56 и 94 определяют факторы
, регулирующие функцию SWR при замене гистона
вариант h3A.Z.
95. Yuan, G.C. etal. Идентификация в масштабе генома
позиций нуклеосом в S.viscerevisiae. Science 309,
626–630 (2005).
96. Маврич, T.N. etal. Модель барьерной нуклеосомы для статистического позиционирования
нуклеосом по всему геному дрожжей
. Genome Res. 18, 1073–1083
(2008).
97. Zofall, M. etal. Гистон h3A.Z взаимодействует с РНКи
.факторов гетерохроматина подавляют антисмысловые РНК.
Nature 461, 419–422 (2009).
98. Zhang, H., Roberts, D.N. И Кэрнс, B.R. Общегеномная динамика
Htz1, варианта гистона h3A, который уравновешивает
репрессированных / базальных промоторов для активации через потерю
гистонов. Cell 123, 219–231 (2005).
99. Dion, M.F. etal. Динамика репликационно-независимого оборота гистонов
у почкующихся дрожжей. Science 315,
1405–1408 (2007).
Ссылки 77 и 99 определяют концепцию, геномное распределение
и факторы, регулирующие оборот гистонов
в дрожжах дикого типа.
100. Durant, M. И Пью, B.F. NuA4-направленный хроматин
транзакций по всему геному Saccharomyces
cerevisiae. Мол. Клетка. Биол. 27, 5327–5335
(2007).
101. Альтаф, М. etal. NuA4-зависимое ацетилирование
нуклеосомных гистонов h5 и h3A непосредственно
стимулирует включение h3A. Z комплексом SWR1
. J.Biol. Chem. 285, 15966–15977
(2010).
102. Halley, J.E., Kaplan, T., Wang, A.Y., Kobor, M.S. &
Райн, J. Роли для h3A. Z и его ацетилирование в транскрипции GAL1
и индукции гена, но не в транскрипционной памяти
GAL1. PLoS Biol. 8, e1000401
(2010).
103. Draker, R. etal. Комбинация h3A.Z и ацетилирования h5
рекрутирует Brd2 на хроматин во время транскрипционной активации
. PLoS Genet. 8, e1003047
(2012).
104. Chen, P. etal.h4.3 активно маркирует энхансеры и активирует транскрипцию гена
посредством открытия более высокого порядка
хроматина. Genes Dev. 27, 2109–2124
(2013).
105. Heinz, S. etal. Выбор и функция клеток
типоспецифических энхансеров. Nature Rev. Mol. Cell Biol.
http://dx.doi.org/10.1038/nrm3949 (2015).
106. Conerly, M.L. etal. Изменения в занятости h3A.Z и метилировании ДНК
во время В-клеточного лимфомагенеза.
Genome Res. 20. С. 1383–1390 (2010).
107. Зильберман, О.Д., Коулман-Дерр, О.Д., Баллинджер, О.Т. &
Henikoff, S. Гистон h3A.Z и метилирование ДНК представляют собой
взаимно антагонистических меток хроматина. Nature 456,
125–129 (2008).
108. Пчелинцев, N.A. etal. Размещение шаперонного комплекса HIRA histone
в хроматиновом ландшафте.
Cell Rep. 3, 1012–1019 (2013).
109. Киреева, M.L. etal. Ремоделирование нуклеосом, индуцированное
РНК-полимеразой II: потеря димера h3A / h3B
во время транскрипции.Мол. Cell 9, 541–552
(2002).
110. Кулаева, О.И. etal. Механизм ремоделирования и восстановления хроматина
при пассаже РНК
полимеразы II. Структура природы. Мол. Биол. 16,
1272–1278 (2009).
111. Белоцерковская, мкр. etal. FACT способствует транскрипционно-зависимому изменению
нуклеосом.
Science 301, 1090–1093 (2003).
ОТЗЫВЫ
ОТЗЫВЫ ПРИРОДЫ
|
ОНЛАЙН-ПУБЛИКАЦИЯ МОЛЕКУЛЯРНОЙ БИОЛОГИИ КЛЕТОК
|
11
В ФОКУСЕ ТРАНСКРИПЦИИ
© Macmillan Publishers Limited, 2015.Все права защищены
Nexiwave — Голосовая почта в текст / Транскрипция голосовой почты
Введение
Как администратор Exchange, вы можете создать правило на стороне сервера Exchange для автоматической пересылки сообщений голосовой почты ваших пользователей в Nexiwave для обработки транскрипции голосовой почты.
Примечание для пользователей Office365: Nexiwave предлагает использовать нашу прямую интеграцию с Office365.
Льготы
- Внешний вид / компьютер пользователей не обязательно должны быть постоянно открытыми.
- Одноразовая конфигурация для всех ваших пользователей.
Отлично подходит для
- Все версии Microsoft Exchange и учетные записи Office365.
Реквизит
- Аккаунт Nexiwave.
- Зарегистрируйте пользователей на странице «Дополнительные учетные записи».
- Ваш исходящий транспорт Exchange позволяет доставлять электронную почту в Nexiwave.
- Ваш входящий транспорт Exchange (и любые спам-фильтры) позволяет доставлять электронную почту от Nexiwave.
- (Примечание: это НЕ работает с Mitel NuPoint Advanced UM. Подробнее см. Ниже.)
Как
( Автоматическая пересылка — одна из самых основных функций в Exchange и поддерживается всеми версиями обмена ВСЕ и Office365. Снимки экрана ниже сделаны с использованием Exchange 2016. Точные шаги очень похожи / идентичны для всех других версий Exchange и Office 365 / Exchange Online )
- Центр администрирования Open Exchange (ECP) для вашей среды Exchange.Вот ссылка на Office365 / Exchange Online.
- Щелкните поток почты на левой панели.
- Перейдите на вкладку rules и щелкните значок «+». Откроется список действий. Нажмите Создать новое правило .
- В окне новое правило настройте параметры правила. Назовите правило как угодно, затем перейдите к разделу Применить это правило, если… и щелкните Отправитель… .
- Выберите адрес электронной почты сервера голосовой почты, для которого вы хотите настроить правило. Выделите его в списке и нажмите кнопку add-> . Если адрес электронной почты вашего сервера голосовой почты отсутствует в списке, введите его в текстовое поле после «Проверить имена» и нажмите «Проверить имена». Затем нажмите OK .
- Щелкните Дополнительные параметры… внизу. Это открывает больше действий.
- ( Необязательно ) Используйте «добавить условие», чтобы при необходимости добавить дополнительную фильтрацию.
Например, во время начального теста попробуйте добавить условие, чтобы это правило сначала применялось только к некоторым тестовым пользователям (или группам).
Примечание: не забудьте удалить это условие после завершения тестирования. - В Выполните следующий раздел , выберите Перенаправить сообщение на действие … .
- В следующем окне введите «[email protected]» в качестве получателя и нажмите «Проверить имена»:
- (Правило в основном гласит: сначала перенаправляйте сообщения голосовой почты, отправленные моим сервером голосовой почты, на Nexiwave)
- Подтвердите свой выбор, нажав кнопку OK .Затем нажмите Сохранить , чтобы завершить настройку правила.
Вот и все — сообщения голосовой почты ваших пользователей теперь будут сначала отправляться в Nexiwave для расшифровки. Nexiwave расшифрует и отправит транскрибированную версию электронного письма (включая стенограмму и аудио) обратно вашему пользователю.
Примечание: после проверки того, что все работает, не забудьте удалить условия тестирования, если они есть, которые вы добавили на шаге 6 выше.
Банкноты
- Включите «Автоматическую регистрацию» или вручную укажите своих пользователей на странице дополнительных учетных записей Nexiwave, в противном случае их сообщения голосовой почты будут игнорироваться.
- Убедитесь, что в вашей установке Exchange есть правило транспорта, разрешающее исходящую доставку и входящее получение от Nexiwave.
- Это НЕ будет работать с функцией Mitel NuPoint Advanced UM. Это связано с тем, что расширенная единая система обмена сообщениями доставляет сообщения через веб-службу Exchange вместо стандартного протокола электронной почты. Вы можете: 1) если вы используете Office365, включить прямую интеграцию Nexiwave O365 для своей учетной записи O365 Enterprise; 2) Или используйте стандартную единую систему обмена сообщениями NuPoint для доставки сообщений через стандартный почтовый протокол.
Вопросы и ответы
- Нужно ли мне менять адрес электронной почты голосовой почты каждого пользователя на собственный адрес электронной почты Nexiwave?
Нет. Когда электронное письмо отправляется на «[email protected]», наша система достаточно умна, чтобы проверять других получателей в заголовках электронной почты и находить их соответствующие учетные записи Nexiwave. - Содержит ли электронное письмо с транскрипцией Nexiwave как стенограмму, так и аудио?
Да, копия Nexiwave всегда содержит: расшифровку стенограммы, аудио-вложение и исходный текст электронного письма (прилагается под расшифровкой).Вся информация переносится. - Могу ли я выбрать только BCC на Nexiwave, кроме перенаправления?
Да, конечно. На шаге 6 выше просто выберите «Добавить получателей» и BCC на «[email protected]». Одна из проблем заключается в том, что ваши пользователи будут получать два сообщения электронной почты с одной и той же голосовой почтой: одно с вашего сервера голосовой почты и одно расшифрованное сообщение от Nexiwave. Выбор за вами.
Быстрый обмен глюкокортикоидного рецептора на промоторе связан с транскрипцией и регулируется шаперонами и протеасомами
На основании восстановления флуоресценции после анализа фотообесцвечивания (FRAP) сейчас известно, что большинство ядерных белков являются очень мобильными (28).Список быстро перемещающихся белков включает факторы сплайсинга (17), ядрышковые белки (9), гистон h2 (19, 23) и несколько факторов транскрипции стероидных рецепторов (8, 22, 31, 34, 43). Во всех случаях подвижность факторов транскрипции медленнее, чем подвижность одного GFP, демонстрируя, что все эти белки временно взаимодействуют с сайтами связывания ядер того или иного типа. Большинство этих сайтов не могут быть специфическими промоторами, учитывая количество экспрессируемых молекул (34). Скорее, связывание с хроматином или ядерным матриксом более вероятно (34, 43).Таким образом, эти ядерные данные FRAP дают представление о перемещении белков внутри ядра, но они не касаются непосредственно связывания фактора транскрипции с промотором.
В ограниченном числе случаев было изучено связывание факторов транскрипции со специфическими промоторами. Здесь снова была обнаружена подвижность, что указывает на то, что факторы транскрипции не остаются постоянно связанными с промотором, а скорее подвергаются циклам связывания и расцепления. Первое доказательство этого было получено в экспериментах с FRAP с использованием тандемного массива промоторных сайтов вируса опухоли молочной железы мышей (MMTV), визуализированных с помощью (GFP) -меченного глюкокортикоидного рецептора (GR) (22).Наблюдался быстрый обмен этого рецептора с общим временем восстановления менее минуты. Более поздние исследования с использованием иммунопреципитации хроматина (ChIP) показали, что рецептор эстрогена (ER) циклически действует на нескольких разных промоторах, но с заметно большей периодичностью, порядка 1 часа (31, 37). В дополнение к самим факторам транскрипции, связанные факторы также демонстрируют обмен на промоторах, но опять же с очень разными временными масштабами в зависимости от того, является ли экспериментальный подход FRAP, где наблюдается быстрый обмен (3, 42), или ChIP, где в целом более медленный обнаружено циклирование (4, 31, 37).Когда временное разрешение ChIP было доведено до предела, реципрокная цикличность двух комплексов коактиватора ER (DRIP и p160) могла быть обнаружена на временной шкале всего 2,5 мин (4). Эти данные предполагают, что в других системах существует гораздо более быстрый обмен, но на уровне или ниже пределов временного разрешения чипа.
Еще многое предстоит узнать о механизмах циклирования транскрипционных факторов. Медленный цикл ER требует протеасомной активности (31). В случае быстрого обмена GR ничего не известно, кроме намеков на участие шаперонов и ремоделеров хроматина.Freeman и Yamamoto (12) показали, что молекулярный шаперон p23 может вызывать разборку транскрипционных регуляторных комплексов тироидных рецепторов. Они также обнаружили, что нацеливание in vivo слитого белка gal4-p23 на промотор GR может значительно снизить там активацию транскрипции. Основываясь на этих и других экспериментах, они предположили, что p23 может участвовать в удалении GR во время быстрого обмена. Система ремоделирования хроматина in vitro показала, что рекрутирование Swi / Snf сопровождается потерей GR, что позволяет предположить, что ремоделеры хроматина могут играть роль в быстром обмене GR (11).
Функция обмена фактора транскрипции также недостаточно изучена. В случаях, выявленных на сегодняшний день, было высказано предположение, что цикл рецепторов на промоторе является механизмом определения изменений в уровнях гормонов (12, 22, 31, 37). Также было высказано предположение, что протеасомное удаление мощных факторов транскрипции с их промоторов может быть средством ограничения транскрипционного выхода (24). Однако, помимо этих гипотез, не было предложено никаких других функций для обмена фактора транскрипции.
Нашей целью в этом исследовании было изучить механизм и функцию быстрого обмена GR, наблюдаемого в живых клетках в тандемном массиве промоторных сайтов MMTV. Мы обнаружили, что в целевых сайтах присутствуют как шапероны, так и протеасомы. Нарушение любого из них приводит к противоположным изменениям обменного курса, указывая на то, что обменный курс обычно частично регулируется балансом между шаперонной и протеасомной активностями. Мы также обнаружили корреляцию между обменом GR и количеством транскрипции, предполагая, что более длительное время пребывания GR способствует большей транскрипции.
МАТЕРИАЛЫ И МЕТОДЫ
Клеточные линии. В большинстве экспериментов использовали клеточную линию 3617. Клетки содержат 200 тандемных повторов элемента размером 9 т.п.н., состоящего из промотора MMTV, за которым следуют ras и гены BPV (16), и они стабильно экспрессируют GFP-меченый GR под контролем системы отключения тетрациклина (44). Контрольные эксперименты были выполнены с родительской клеточной линией 3134, которая содержит тандемные повторы, но не GR с меткой GFP. Для создания клеток, содержащих только GFP, 3134 клетки трансфицировали плазмидой GFP (pEGFPC1; Clontech).Для создания клеток, содержащих GFP-HP1α, 3134 клетки трансфицировали плазмидой для конструкции (любезный подарок T. Cheutin и T. Misteli, Лаборатория рецепторной биологии и экспрессии генов, Национальный институт рака, Бетесда, Мэриленд).
Клетки выращивали при 37 ° C с 5% CO. 2 в модифицированной Дульбекко среде Игла (DMEM) (Gibco BRL) с добавлением 2 мМ глутамина (Gibco BRL), 10% фетальной бычьей сыворотки (HyClone) и 5 мкг тетрациклина / мл (для подавления экспрессии GFP-GR). При подготовке к экспериментам под микроскопом клетки переносили в эту среду и оставляли там на ночь, за исключением того, что тетрациклин не добавляли, фетальную бычью сыворотку обрабатывали древесным углем и декстраном (Gemini Bio-Product) для удаления стероидов, которые могли активировать GFP-GR, и Для устранения автофлуоресценции использовали среду, не содержащую фенолового красного.Непосредственно перед экспериментами GFP-GR активировался либо синтетическим гормоном дексаметазоном (Sigma), либо естественным гормоном кортикостероном (Sigma), либо антагонистом RU486 (любезный подарок Кэти Смит, Лаборатория биологии рецепторов и экспрессии генов), каждый в концентрации 100 нМ.
FRAP. Для экспериментов с FRAP клетки выращивали в течение ночи в камерах с покровным стеклом (Lab-Tech), а затем индуцировали гормоном в течение 30 мин. Для ингибирования hsp90 или протеасом после гормональной индукции применяли гельданамицин (2,5 мкг / мл) или MG-132 (100 мкМ) (Calbiochem) соответственно.В обоих случаях были проведены параллельные контрольные эксперименты с носителем (диметилсульфоксид). После обработки гелданамицином камеры с живыми клетками держали на предметном столике микроскопа не более 5 минут с кортикостероном или 30 минут с дексаметазоном, чтобы избежать стресса, приводящего к образованию агрегатов GFP-GR. Для экспериментов по истощению АТФ клетки обрабатывали 10 мМ азида натрия (Sigma) в DMEM без глюкозы с добавлением 6 мМ 2-дезоксиглюкозы (Sigma) или с DMEM без глюкозы без азида натрия и только с 6 мМ 2-дезоксиглюкозы.Эксперименты FRAP проводились на конфокальном микроскопе Zeiss 510 либо с числовой апертурой 40 × 1,3, либо с числовой апертурой 100 × 1,3. масляный иммерсионный объектив. Клетки поддерживали при 37 ° C, используя инкубатор с воздушным потоком (Nevtek). Отбеливание проводилось с помощью линий 488 и 514 нм аргонового лазера мощностью 45 мВт, работающего при мощности лазера 75%. Для отбеливающего импульса использовалась одна итерация, которая длилась 0,018 с для объектива 40x или 0,065 с для объектива 100x. Восстановление флуоресценции контролировали при низкой интенсивности лазера (0.2% для лазера мощностью 45 мВт) с интервалами от 40 до 500 мс, в зависимости от эксперимента.
Анализ FRAP. Было выполнено около 10 отдельных FRAP, которые затем усреднились для создания единой кривой FRAP. Временное разрешение оставалось постоянным, пока измерялось восстановление, но это привело к очень большому количеству близко расположенных точек на второй, более медленной фазе кривой восстановления. Чтобы решить эту проблему, мы усреднили от 3 до 10 соседних точек в этой более медленной части кривой. Это привело к созданию примерно одинаково расположенных точек вдоль кривой восстановления и, следовательно, позволило избежать чрезмерного взвешивания более медленной фазы кривой во время подгонки.
Эффективная диффузионная подгонка была выполнена с использованием теории Soumpasis для круглого пятна отбеливания (39), реализованной в Matlab с помощью программы nlinfit. В теории Soumpasis скорость FRAP определяется как τ = w 2 /4 D , где w — радиус пятна отбеливания, а D — константа диффузии (или эффективной диффузии). Данные FRAP представлены как frap ( t ) = e −2 r / t [ I 0 (2 r / t ) + I 1 (2 r / t )], где t — время, а I 0 и I 1 — модифицированные функции Бесселя.Функции Бесселя и их варианты обычно возникают в дифференциальных уравнениях с цилиндрической симметрией (1), как это происходит для FRAP с круглым пятном отбеливания (15). Теория Soumpasis предполагает нормализованный FRAP, который находится в диапазоне от 0 до 1. Чтобы учесть это, мы перенормировали наши данные FRAP, установив глубину отбеливания на 0, а окончательный уровень восстановления на 1. Это исключает любые различия между восстановлением из-за отбеливателя. глубина или конечная неподвижная фракция. Таким образом, параметр τ напрямую измеряет скорость подъема кривой восстановления.Для каждого FRAP оценка τ дала стандартную ошибку, которая затем использовалась в тесте t для вычисления значения P при статистическом сравнении двух кривых восстановления. Эти значения P показаны на всех сравнительных графиках для извлечения GFP-GR.
Прогнозируемая константа диффузии может использоваться для оценки массы молекулы при условии отсутствия связывающих взаимодействий. Оценка Soumpasis для ядерной подвижности GFP-GR дает оценку D GFP-GR из 1.05 мкм 2 / с. Soumpasis, пригодный для GFP только в ядрах той же клеточной линии, дает оценку D GFP 15 мкм 2 / с (данные не показаны), в ~ 15 раз медленнее, чем GFP-GR. С D ∝ M -1/3 , где M — масса, данные FRAP предсказывают увеличение массы GFP-GR на 15 3 по сравнению с GFP, или молекулярную массу на ~ 95 МДа для GFP-GR. Это намного больше, чем любой известный молекулярный комплекс, и поэтому подвижность GFP-GR в нуклеоплазме должна быть замедлена за счет связывающих взаимодействий.{{\ ast}} _ {on} / \ mathit {k} _ {off} \) $$ mathtex $$ . Тогда k от изменяли, как указано в легенде к рисунку, чтобы получить новые значения для τ, которые затем использовались для построения новых кривых FRAP из уравнения Soumpasis, описывающего FRAP.
РИС. 1.FRAP в массиве MMTV и в других частях ядра зависят от размера пятна обесцвечивания и хорошо соответствуют модели эффективной диффузии. (а) Клетка 3617 после 30-минутной индукции 100 нМ дексаметазона. Массив MMTV виден как яркое пятно (обведено) возле одного из ядрышек.Шкала 10 мкм. (b) Для быстрого сбора данных во время FRAP изображения были собраны только в полосе, охваченной красным прямоугольником на панели a. Отображаются выбранные моменты времени (t). (c) Матрицы были обесцвечены с радиусом пятна 0,9 или 1,8 мкм. Во всех случаях массив был достаточно большим, чтобы полностью заполнить область пятна отбеливания. Более крупные пятна отбеливания приводят к значительно более медленному выздоровлению. Это указывает на то, что диффузия способствует FRAP. Для этих и всех последующих кривых FRAP стандартные ошибки для всех точек всегда были <0.01 и, следовательно, меньше, чем точки, используемые для построения. (d) Восстановление GFP-GR в другом месте ядра также показывает зависимость от размера пятна обесцвечивания, что указывает на роль диффузии GFP-GR в подвижности ядра. (e) Данные GFP-GR FRAP как в массиве, так и в другом месте ядра хорошо соответствовали однопараметрической модели для диффундирующей молекулы, обесцвеченной круглым пятном, со скоростью восстановления, задаваемой параметром подгонки τ. Восстановление в массиве постоянно происходит медленнее, чем где-либо в ядре.Эта разница статистически значима, как показывает тест t с использованием средних значений τ и их 95% доверительных интервалов. Вычисленное значение P (<0,0001) очень важно. (f) Модели FRAP, которые включают как диффузию, так и связывание, показывают, что значительные изменения параметров связывания приводят к небольшим изменениям в кривой FRAP. Это связано с тем, что диффузия остается неизменной после обработок, влияющих на связывание. Прогнозируемые кривые показаны для ряда отклонений.
Анализ транскрипции. Уровни транскрипции измеряли в матрице MMTV с помощью флуоресцентной гибридизации РНК in situ (FISH) точно так, как описано Müller et al. (25). Конкретные условия для каждого транскрипционного анализа были следующими. При изучении эффектов гелданамицина клетки индуцировали гормоном в течение 15 минут, чтобы позволить GFP-GR переместиться в ядро, а затем обрабатывали в течение 45 минут лекарственным средством или носителем (диметилсульфоксид). Чтобы определить уровни транскрипции после лечения препаратом, уровни РНК FISH корректировали с учетом транскрипции в первые 15 мин, когда присутствовал гормон, но до добавления какого-либо лекарства.Это было сделано путем измерения средних уровней транскрипции через 15 минут гормонального лечения и последующего вычитания этого значения из средней интенсивности РНК FISH после 45-минутного лечения лекарственными препаратами.
Для MG-132 клетки предварительно обрабатывали препаратом в течение 1,5 ч и индуцировали гормоном в течение 30 мин перед фиксацией. Во всех случаях средние уровни транскрипции были получены по крайней мере из 35 клеток. Когда измеряли уровни транскрипции конденсированных и деконденсированных массивов, добавляли 100 нМ дексаметазона, а затем через 3 часа клетки готовили для РНК FISH.Всего было исследовано 113 ячеек, и периметр диаметром 6 мкм был выбран в качестве порогового значения, отделяющего сконденсированные массивы от деконденсированных.
Иммунофлуоресценция. Клетки выращивали в течение ночи на 22-миллиметровых покровных стеклах 2 , затем индуцировали 100 нМ дексаметазоном или кортикостероном и фиксировали абсолютным метанолом при -20 o C в течение 15 минут или в течение 5 минут в 1 части 37%. формальдегид и 9 частей буфера PEM (100 мМ PIPES, 5 мМ EGTA, 2 мМ MgCl 2 , pH 6,8, плюс 0.2% Тритон Х-100). После любой фиксации клетки трижды промывали фосфатно-солевым буфером (PBS) по 10 мин каждый раз. Сходные результаты иммунофлуоресценции были получены с обоими протоколами фиксации, за исключением того, что фиксация метанолом требовалась для обнаружения любого окрашивания антителом hsp90.
Для обнаружения агрегатов GFP-GR индуцированные кортикостероном клетки обрабатывали в течение 10 мин 2,5 мкг гелденамицина / мл с последующими 20 мин тепловым (45 ° C) или холодным (на льду) шоком и фиксировали в 3 раза. .5% параформальдегид при комнатной температуре в течение 15 мин. После фиксации формальдегидом клетки подвергали проницаемости в течение 10 минут с помощью 0,5% Triton X-100 в PBS и трижды промывали в PBS по 10 минут каждый раз.
Для всех экспериментов по иммунофлуоресценции клетки инкубировали в течение ночи с первичным антителом, разведенным в PBS с 4% бычьим сывороточным альбумином и 0,1% Tween 20. После инкубации клетки промывали три раза в PBS по 10 мин каждый раз, а затем инкубировали в течение 1-2 часа с соответствующим вторичным антителом, конъюгированным либо с техасским красным, либо с родамином.Затем клетки промывали еще три раза в PBS перед окончательной фиксацией в PBS и исследовали на микроскопе Leica DMRA с Leica 100 × 1,3-N.A. масляный иммерсионный объектив. Изображения были получены в зеленой (GFP-GR) и красной (антитела) флуоресценции с помощью камеры SenSys (Photometrics) с чипом KAF1400, сконфигурированным для сбора пикселей диаметром 0,067 мкм.
Были использованы следующие антитела: анти-hsp70 (MA3-006; аффинные биореагенты), анти-p23 (MA3-414; аффинные биореагенты), антипротеасома (19S субъединица S10B; аффинные биореагенты) и анти-hsp90 (SPA-835 ; Стрессген).
РЕЗУЛЬТАТЫ
Характеристика кривых FRAP. Для исследования механизма и функции быстрого обмена фактора транскрипции мы использовали линию клеток 3617, в которой обмен GR был ранее охарактеризован (22). Эти клетки представляют собой аденокарциномы мыши, содержащие ~ 200 тандемных копий промотора MMTV с ассоциированными репортерными генами (44). Путем количественной оценки уровней GFP в массиве MMTV мы подсчитали, что в среднем там присутствует ~ 700 GFP-GR, что согласуется с ожидаемым количеством сайтов связывания GR в массиве (10).
В клетках, обработанных гормоном, массив MMTV становится видимым как яркое пятно (25) (рис. 1a). FRAP на этом участке показывают очень быстрое восстановление с половиной времени (время до 50% восстановления) ~ 5 с (22). Для достижения хорошего временного разрешения этого восстановления данные FRAP собирались с интервалами ≥40 мс путем сбора изображений только в узкой полосе, охватывающей массив (рис. 1a и b). Приблизительно 10 клеток были усреднены для создания одной кривой FRAP для одного эксперимента, и все эксперименты, описанные здесь, были выполнены, по крайней мере, в три разных дня, чтобы гарантировать воспроизводимость.
FRAP измеряет подвижность за счет диффузии и, если присутствует, транспорта или связывания. Распространение способствует каждому выздоровлению, но в некоторых случаях этот вклад невелик и / или быстр, и его можно игнорировать. Чтобы проверить роль диффузии, мы выполнили отбеливание с различными размерами пятен на массиве и наблюдали различия в восстановлении (рис. 1c). Мы также обнаружили различия в скорости восстановления, когда другие области ядра были обесцвечены с помощью пятен разного размера, что указывает на то, что диффузия способствовала восстановлению GFP-GR повсюду в ядре (рис.1г).
Эти скорости восстановления, однако, были значительно ниже, чем у одного GFP, который восстанавливается почти мгновенно с размерами пятна, используемыми в наших экспериментах (данные не показаны). Если бы восстановление GFP-GR происходило за счет простой диффузии, то они предсказывали бы массу GFP-GR ~ 95 МДа, что значительно больше, чем у любого известного комплекса (подробности этого расчета см. В Материалах и Методах). Следовательно, связывающие взаимодействия должны задерживать GFP-GR FRAP как в ядре, так и особенно в массиве MMTV.
Самый простой сценарий для объяснения комбинированной диффузии и связывания — это эффективная диффузия (15). Здесь FRAP демонстрирует замедленную диффузию с «эффективной» константой диффузии, замедленной пропорционально аффинности связывания. В соответствии с эффективной диффузией, скорости GFP-GR FRAP хорошо соответствовали однопараметрической модели для диффундирующей молекулы, обесцвеченной круглым пятном (рис. 1e) (39).
Подгонки эффективной диффузии информативны по нескольким причинам. Во-первых, они обеспечивают однопараметрическую аппроксимацию скорости FRAP (τ), что позволяет проводить статистическое сравнение кривых на основе этого параметра.Например, при использовании того же размера пятна отбеливания τ в массиве значительно больше, чем τ в другом месте ядра (рис. 1e). Эти значения τ с их 95% доверительным интервалом могут использоваться в тесте t для вычисления значения P , чтобы оценить, существенно ли отличаются восстановления. Значение P для извлечения ядра по сравнению с извлечением массива очень важно ( P <0,0001). Это демонстрирует, что массив восстанавливается значительно медленнее, чем другие сайты в ядре, что указывает на более плотное связывание в массиве, как и ожидалось для локальной высокой концентрации там специфических сайтов связывания.
Во-вторых, теория эффективной диффузии дает представление о том, как такие кривые FRAP изменяются в ответ на изменения аффинности связывания. Удвоение или уменьшение вдвое скорости восстановления приводит к умеренным изменениям скорости восстановления, поскольку диффузия, которая не изменяется, также вносит свой вклад (рис. 1f). Таким образом, в сценарии эффективной диффузии данные FRAP должны быть собраны и проанализированы с точностью, чтобы обнаружить изменения в аффинности связывания при различных возмущениях.
Наконец, в своей простейшей форме теория эффективной диффузии предсказывает константу псевдоравновесного связывания (15), но только для случая, когда распределение флуоресценции является однородным.Это должно относиться к типичным местоположениям в ядре, но в настоящее время такие оценки не могут быть получены для массива, потому что теория должна быть расширена для учета такого неоднородного распределения флуоресценции, в котором яркая область (массив) окружена более тусклой областью. (остальное ядро). Дальнейшие теоретические исследования необходимы для извлечения оценок констант связывания из этих данных.
Во всех последующих экспериментах мы использовали оценки скорости извлечения τ на установке только как средство для сравнения кривых.Для любой пары восстановлений GFP-GR мы выполнили тест t для значений τ для скорости восстановления. На каждом графике мы сообщаем значение P как меру того, существует ли статистически значимая разница между кривыми.
Энергетическая зависимость восстановления GFP-GR. Простейшее объяснение обмена GFP-GR в массиве состоит в том, что он отражает пассивный процесс, определяемый простым равновесием связывания GFP-GR с промотором MMTV. Если это так, то процесс обмена должен быть энергетически независимым.Чтобы проверить это, мы истощили клеточные уровни АТФ с помощью азида натрия и дезоксиглюкозы. FRAPs как в массиве, так и в другом месте ядра были радикально изменены (рис. 2a и b). Оба участка показали восстановление <100% с более значительным замедлением в массиве MMTV, чем где-либо еще в ядре. Это предполагает, что для нормального обмена GFP-GR в массиве может потребоваться энергия, но большое влияние на восстановление в другом месте ядра вызвало беспокойство о неспецифических эффектах. Поэтому мы стремились определить условия для восстановления АТФ, которые минимизируют влияние на FRAP в других сайтах ядра.Это было достигнуто путем инкубации только с дезоксиглюкозой, которая блокирует только неокислительное фосфорилирование (36) и поэтому не так эффективна, как азид и дезоксиглюкоза вместе. FRAP были слегка затронуты во всем ядре после обработки дезоксиглюкозой, но восстановление в матрице теперь было качественно другим, поскольку они все еще демонстрировали неподвижную фракцию (рис. 2c и d). Эта неподвижная фракция исчезла после вымывания дезоксиглюкозы, демонстрируя, что неподвижность не была следствием токсичности (рис.2д).
РИС. 2. ОбменGFP-GR в массиве является энергозависимым процессом. 30-минутная обработка 10 мМ азида натрия и дезоксиглюкозы приводит к заметному снижению скорости обмена в массиве (а) и в других местах ядра (б). Перенос клеток в среду без глюкозы (только дезоксиглюкозу) на 60-90 мин индуцирует около 5% неподвижной фракции в массиве (c), но обмен на других участках ядра не показывает неподвижную фракцию (d) . (e) Вымывание дезоксиглюкозы устраняет неподвижную фракцию в матрице.
Полное восстановление, наблюдаемое в другом месте ядра, предполагает, что пониженные уровни АТФ сначала влияют на подмножество сайтов, уникальных для массива MMTV, прежде чем повлиять на связывание общего GR-хроматина. Эта часть сайтов MMTV, по-видимому, требует энергии для выпуска GFP-GR. Однако быстрый обмен все еще происходит в оставшихся сайтах MMTV, тем самым определяя два класса сайтов: небольшая фракция, чувствительная к АТФ, и большая часть, которая относительно нечувствительна к АТФ. Эти результаты резко контрастируют со многими ядерными белками, которым не требуется энергия для своей подвижности (19, 28).
Роль шаперонов в восстановлении GFP-GR. Ряд факторов может вносить вклад в энергетическую зависимость АТФ-чувствительной фракции сайтов MMTV. АТФ-зависимый шаперонный комплекс, который включает hsp70, hsp90, p23 и несколько иммунофилинов, связывается с не связанными границами GR в цитоплазме, обеспечивая связывание лиганда (30). Как только лиганд связывается, шаперонный комплекс диссоциирует, но позже воссоединяется после того, как лиганд отделяется от GR. Эта ассоциация и диссоциация, вызванные потерей лиганда, называются шаперонным циклом.Помимо своей цитоплазматической роли, шаперонный комплекс также играет роль в рециклинге GR в ядре (7, 20). Более того, Freeman и Yamamoto (12) предположили, что шапероны разбирают транскрипционные регуляторные комплексы на промоторных сайтах. Если это имело место для GR в промоторе MMTV, то нарушение функции шаперона должно привести к замедлению FRAP в массиве MMTV.
В качестве первого теста на участие шаперонов мы использовали иммунофлуоресценцию для исследования субклеточного распределения hsp90, hsp70 и p23 в клеточной линии массива MMTV.Мы обнаружили, как и ожидалось, что эти молекулы были в основном цитоплазматическими, но также была обнаружена некоторая ядерная флуоресценция. Примечательно, что мы последовательно обнаружили ассоциацию hsp90, hsp70 и p23 с массивом MMTV. Антитела против этих молекул совместно локализовались с сигналом GFP-GR в матрице, а также обычно окрашивали область, окружающую матрицу (рис. 3). Размер области, окрашенной шаперонами, всегда был пропорционален размеру массива MMTV, с более крупными массивами, которые содержали больше GFP-GR, характеризуемых, соответственно, более крупными областями окрашивания шаперонов.Эти наблюдения демонстрируют, что шапероны рекрутируются на сайт MMTV, и предполагают, что шапероны способны влиять на обмен GFP-GR в массиве.
РИС. 3.Иммунофлуоресцентное обнаружение белков-шаперонов на матрице MMTV (обведено). Антитела против hsp90 (от a до c), hsp70 (от d до f) или p23 (от g до i) окрашивают цитоплазму, как и ожидалось, но внутри ядер они также последовательно колокализуются с GFP-GR в массиве MMTV. Вставки содержат увеличенные изображения колокализации в массиве.Шкала 10 мкм.
Чтобы нарушить активность шаперона, мы сначала использовали антибиотик гелданамицин (32), который специфически блокирует активность hsp90, связываясь с его N-концевым сайтом АТФ и предотвращая связывание p23 (38). Как и ожидалось, гелданамицин (2,5 мкг / мл) блокировал импорт GFP-GR в клеточной линии массива (6), поэтому для изучения ядерных событий мы предварительно обработали клетки дексаметазоном, а затем добавили гелданамицин через 30 минут, когда ядерный импорт GR был завершен. Хотя массивы MMTV начали исчезать через 1 час лечения гелданамицином (рис.4a), нормальное количество массивов все еще было ясно видно после 30-60 мин обработки гелданамицином, что позволяло измерять с помощью FRAP обмен GFP-GR в матрице. В клетках, обработанных гелданамицином, FRAP в матрице были быстрее, чем в контрольных клетках (фиг. 5a). Более сильные эффекты можно было наблюдать при более высоких концентрациях гелданамицина, но они также приводили к значительным эффектам в других частях ядра (данные не показаны). Таким образом, используемая здесь концентрация гелданамицина сводит к минимуму воздействие на остальную часть ядра и, следовательно, должна отражать эффекты, уникальные для связывания GFP-GR с сайтами MMTV.Дальнейшие доказательства этого получены из клеток, трансфицированных другим ядерным белком, GFP-HP1α, и обработанных гелданамицином. Никакого влияния на восстановление GFP-HP1α обнаружено не было (рис. 5b). Аналогичные результаты были получены и для одного GFP (данные не показаны). Эти данные показывают, что гелданамицин в целом не изменяет подвижность белков в ядре.
РИС. 4.Массивы (обведены) исчезают намного быстрее после лечения гелданамицином, когда кортикостерон является лигандом, чем когда дексаметазон является лигандом.В любом случае гельданамицин вызывает исчезновение массивов (красные линии) быстрее, чем обычно (черные линии). Пример исчезновения массива для каждого случая показан псевдоцветом, чтобы выделить массивы. (а) Когда дексаметазон является лигандом, исчезновение массива происходит постепенно и происходит в течение 2 часов. (b) Когда кортикостерон является лигандом, исчезновение матриц, вызванное гелданамицином, резко ускоряется и происходит в течение 10 минут. Эти результаты показывают, что эффекты гелданамицина усугубляются кортикостероном, предположительно из-за его более быстрого обмена с ГР.Они также предполагают, что в присутствии гелданамицина GR в конечном итоге становится лишенным лиганда и неспособным связываться с сайтами MMTV. Шкала 5 мкм.
РИС. 5.Эффекты ингибирования hsp90. FRAP в массивах MMTV быстрее после обработки 2,5 мкг гелданамицина / мл (a) или 5 мкг радицикола / мл (c), чем в контрольных клетках, но ускорение обнаруживается сразу в кортикостероне (e), пока оно появляется только через 30-60 мин в дексаметазоне (а). (b) Эти изменения не вызваны общим влиянием на подвижность белка, поскольку клетки, трансфицированные GFP-HP1α, не проявляют никакого эффекта на FRAP после обработки гелданамицином.(Обратите внимание, что извлечение GFP-HP1α не соответствует эффективной диффузии [данные не показаны], поэтому тест t для вычисления значения P не может быть проведен.) (D) Эксперименты по отмене гормона демонстрируют это по сравнению с дексаметазоном. , кортикостерон намного быстрее обменивается с ГР. Показаны средние уровни транскрипции, измеренные с помощью РНК FISH для 35 клеток. Клетки индуцировали 100 нМ кортикостероном (CORT) или дексаметазоном (DEX) в течение 15 минут, а затем трижды промывали в течение 5-минутного периода в среде без гормонов и оставляли в этой среде на 45 минут.При использовании кортикостерона в качестве лиганда транскрипция отменяется после 5-минутной промывки, что указывает на полную замену лиганда на GR во время промывки. В тот же период отмывки значительное количество дексаметазона остается связанным, поскольку транскрипция снижается только на 50%. (f) Обработка гелданамицином вызывает прогрессирующую потерю GFP-GR из массива и уменьшение размера, что сопровождается потерей шаперонов. Показана потеря p23 из массива после обработки гелданамицином (GELD) дексаметазоном в качестве лиганда.Шкала 5 мкм.
Чтобы подтвердить результаты гелданамицина, мы также обрабатывали клетки другим препаратом, радициколом, который нарушает активность hsp90, блокируя его N-концевой сайт связывания АТФ (35). И снова мы обнаружили, что FRAP в массиве был быстрее (рис. 5c).
Известно, что гелданамицин и радицикол специфичны для hsp90 (32), но этот шаперон играет роль во многих клеточных процессах (29). Чтобы проверить, влияет ли гелданамицин напрямую на ГР, мы заменили дексаметазон естественным гормоном кортикостероном.Поскольку полупериод связывания кортикостероидов с GR при 0 ° C намного короче, чем у дексаметазона (14, 26), мы пришли к выводу, что кортикостерон должен усиливать любые эффекты гелданамицина, если они возникают из-за потери лиганда из GR и последующего прохождения через шаперонный цикл.
Чтобы подтвердить разницу в аффинности связывания этих двух стероидов с GFP-GR в клеточной линии массива MMTV при физиологических температурах, мы использовали кортикостерон или дексаметазон в качестве лиганда, промыли клетки три раза в течение 5-минутного периода в среда без лиганда, и сравнили транскрипционные выходы.После вымывания кортикостерона дополнительная транскрипция была отменена, в отличие от вымывания дексаметазоном, где транскрипция упала только на 50% (рис. 5d). Это демонстрирует, что в течение 5-минутного периода отмывки практически весь кортикостерон стал несвязанным с GR, тогда как значительные количества дексаметазона все еще были связаны. Эти результаты демонстрируют in vivo, что дексаметазон остается гораздо более тесно связанным с GR, чем кортикостерон.
Как и предполагалось, эффекты гелданамицина усиливались в присутствии кортикостерона.Исчезновение массивов GFP-GR происходило намного быстрее с кортикостероном (фиг. 4b), а FRAP становилось быстрее сразу после добавления гелданамицина (фиг. 5e). Этот немедленный ответ убедительно свидетельствует о том, что эффект гелданамицина специфичен для GR.
Затем мы спросили, влияет ли лечение гелданамицином на ассоциацию шаперонов с массивом MMTV. Используя иммунофлуоресценцию, мы обнаружили, что уровни p23 снижались пропорционально продолжительности лечения гелданамицином (рис. 5f).Аналогичные результаты наблюдались с hsp90 и hsp70. Постепенная потеря шаперонов всегда сопровождалась уменьшением размера массива и количества ассоциированного GFP-GR. Таким образом, наши данные не различают, приводит ли потеря шаперонов после лечения гелданамицином к потере GFP-GR или наоборот.
Более быстрые FRAP в присутствии гелданамицина также наблюдались после более продолжительного лечения (от 1 до 3 часов с дексаметазоном в качестве лиганда), хотя массивы стали труднее обнаруживать (рис.4а). После 30 мин визуализации этих клеток, обработанных гелданамицином в течение 1–3 ч, по всему ядру появились многочисленные пятна, обогащенные GFP-GR (рис. 6а). Эти пятна не были массивами, поскольку в ядре присутствуют только один или два массива. Пятна также могут возникать, когда лечение гелданамицином сочетается с холодом или тепловым шоком. Пятна совместно локализованы с протеасомным компонентом (Рис. 6b и c), указывая тем самым, что они могут отражать аномальные скопления неправильно свернутых белков (41). Как наблюдалось для этих других включений, связанных с протеасомами, FRAP в пятнах GFP-GR обнаруживали замедление с неподвижной фракцией (рис.6г). Это указывает на то, что некоторая фракция GR иммобилизуется в каждом пятне и образует белковый агрегат. Таким образом, мы обнаружили, что продолжительное лечение гелданамицином в сочетании с дополнительным стрессом (тепло, холод или визуализация) может вызывать неспецифические эффекты, которые приводят к более медленному FRAP на агрегатах GFP-GR. Как только эти агрегаты сформировались, массивы исчезли, но до этого момента FRAP в массивах всегда был быстрее, чем в элементах управления.
РИС. 6.Ядерные агрегаты GFP-GR. (от a до c) Обработка гелданамицином в сочетании со стрессом (тепло, холод или длительная визуализация) вызывает исчезновение массивов и образование пятен GFP-GR, которые совместно локализуются с протеасомными антителами.(d) В этих пятнах иммобилизована часть GFP-GR по сравнению с другими областями ядра. Во время визуализации эти агрегаты появляются быстрее с кортикостероном, чем с дексаметазоном. Шкала 5 мкм.
В целом гелданамицин или радицикол вызывали более быстрое выздоровление в матрице. Это ускорение происходило мгновенно в присутствии кортикостерона, что убедительно свидетельствует о специфическом влиянии на GFP-GR. Следовательно, неподвижная фракция, наблюдаемая после истощения АТФ, не является результатом разрушения шаперонов.
Роль протеасомы в восстановлении GFP-GR. Протеасома является альтернативным кандидатом для регуляции АТФ-чувствительной фракции сайтов MMTV. И 19S, и 20S субъединицы протеасомы нуждаются в АТФ (18). Как отмечалось во введении, все больше данных указывает на роль протеасомы в регуляции факторов транскрипции, и, более того, протеасомы часто взаимодействуют с шаперонами.
В качестве первого теста на участие протеасом мы окрашивали клетки антителом против компонента 19S.Это антитело последовательно колокализовалось с массивом и окружающими областями, предполагая, что протеасома рекрутировалась в этот сайт и могла играть роль в обмене GFP-GR там (рис. 7a-c). Как было обнаружено для компонентов шаперонов, существует прямая корреляция между размером массива и степенью окрашивания протеасом (данные не показаны), предполагая, что увеличение количества GR привлекает больше протеасом к промотору.
РИС. 7.Иммунофлуоресцентное обнаружение протеасомы в массиве MMTV (обведено) и FRAP после нарушения функции протеасомы.Обнаружено значительное окрашивание протеасом в цитоплазме. (от a до c) Внутри ядра постоянно наблюдалась совместная локализация с массивом MMTV. На вставке показан увеличенный вид колокализации в массиве. (d и e) Клетки, индуцированные дексаметазоном или кортикостероном и подвергшиеся действию ингибитора протеасом MG-132, проявляли более медленную FRAP в массивах MMTV. (f) Это различие не было вызвано общим замедлением подвижности белков в ядре, поскольку обработка MG-132 не влияла на восстановление GFP-HP1α.(Опять же, эти извлечения GFP-HP1α не соответствуют эффективной диффузии, поэтому ни скорость извлечения, τ, ни значение P для сравнения не могут быть рассчитаны.) Масштабная шкала, 10 мкм.
Чтобы проверить роль протеасомы в обмене GFP-GR в массиве MMTV, мы обработали клетки ингибитором протеасом MG-132 (18). Это привело к замедлению FRAPs с дексаметазоном или кортикостероном в качестве лиганда (рис. 7d и e). Более сильные эффекты с MG-132 можно было вызвать при более высоких концентрациях или более длительном лечении, но мы выбрали концентрацию и время лечения так, чтобы эффекты MG-132 были минимизированы в другом месте ядра.Это должно помочь гарантировать, что эффекты, наблюдаемые в массиве MMTV, были уникальными для привязки GFP-GR к сайтам MMTV. Кроме того, MG-132 не влиял на подвижность только GFP (данные не показаны) или GFP-HP1α (фиг. 7f), демонстрируя, что он не нарушал неспецифически подвижность белка по всему ядру.
Имея доказательства активности протеасом и шаперонов на сайтах MMTV, мы затем спросили, повлияло ли нарушение одной системы на другую. Мы использовали иммунофлуоресценцию для изучения уровней протеасомы 19S после лечения гелданамицином.Как было замечено ранее для компонентов шаперона, гелданамицин также приводил к прогрессирующей потере со временем протеасомы 19S из массива MMTV (фиг. 8a-f). Это всегда коррелировало с уменьшением размера самого массива и количества GFP-GR в нем, что делало невозможным различить, вызывает ли потеря GFP-GR протеасомную потерю или наоборот. Тем не менее, эта постепенная потеря протеасомы после лечения гелданамицином должна сама по себе приводить к более медленным FRAP, тем самым противодействуя обнаруженному нами эффекту, а именно более быстрым FRAP.Это говорит о том, что мы могли наблюдать даже более быстрое выздоровление после того, как гелданамицин не повлиял на уровни протеасом. В дополнительных экспериментах мы также использовали иммунофлуоресценцию для изучения уровней p23 после обработки MG-132 и не обнаружили изменений (рис. 8g-l). Этот эффект был замечен либо с дексаметазоном, либо с кортикостероном, хотя эти разные лиганды по-разному влияли на транскрипцию (см. Ниже). Мы пришли к выводу, что замедление, наблюдаемое после лечения MG-132, не связано с потерей p23.
РИС. 8.Влияние лечения гелданамицином на протеасомы или лечения MG-132 на шапероны. (от a до f) Прогрессирующая потеря протеасомы 19S наблюдается на матрице MMTV (обведено кружком) при более длительной обработке гелданамицином (GELD). Четкое протеасомное окрашивание наблюдается через 15 мин обработки гелданамицином (b), тогда как более слабое окрашивание на уровнях, близких к ядерному фону, обнаруживается через 1 час обработки гелданамицином (e). (от g до l) MG-132 не влияет на уровни p23 ни с дексаметазоном (DEX) (от g до i), ни с кортикостероном (CORT) (от j до l).Для каждого лиганда показана только одна временная точка, поскольку, в отличие от обработки гелданамицином, нет потери GFP-GR из массива после ингибирования протеасом. Шкала 5 мкм.
В целом, наши результаты предполагают, что протеасома сама по себе обычно регулирует обмен GFP-GR на сайтах MMTV и отвечает, по крайней мере, за некоторую чувствительность к АТФ на части этих сайтов.
Уровень транскрипции и восстановление GFP-GR. Предыдущие эксперименты продемонстрировали, что нормальное восстановление GFP-GR зависит от энергии и требует функций протеасомы и шаперона.Это показывает, что обмен GFP-GR строго регулируется. Поскольку транскрипция является основной функцией связывания GR с промотором MMTV, мы спросили, существует ли какая-либо связь между транскрипцией и скоростью обмена.
Для начала мы исследовали, коррелирует ли скорость обмена с различными уровнями эндогенной транскрипции. Наши предыдущие исследования показали, что размер массива коррелирует с уровнем транскрипции, измеренным с помощью RNA FISH. Конденсированные массивы менее транскрипционно активны, чем деконденсированные (25) (рис.9а и таблица 1). Поэтому мы выполнили FRAP на конденсированных и деконденсированных массивах и обнаружили, что восстановление на конденсированных массивах было немного, но стабильно, быстрее, чем на деконденсированных массивах (рис. 9b). Хотя эта разница незначительна, эта разница дает очень значимое значение P ( P <0,001), потому что данные настолько хорошо соответствуют эффективной диффузии, что оценка скорости восстановления для каждой кривой имеет очень небольшую ошибку.
РИС. 9.Ускоренный обмен GFP-GR связан с меньшей транскрипцией.(а) Примеры конденсированного массива и деконденсированного массива показаны с соответствующими сигналами РНК FISH. (б) FRAP конденсированных массивов последовательно выполнялись быстрее. (c и d) RU486 (100 нМ) и кортикостерон (100 нМ) также давали более быстрые FRAP, чем дексаметазон. Масштабная линейка, 1 мкм.
ТАБЛИЦА 1.Изменения в восстановлении GFP-GR в массиве связаны с транскрипцией a
Для дальнейшего исследования связи между скоростью обмена и транскрипцией мы измерили FRAP в присутствии антагониста RU486.RU486 снижал транскрипцию в массиве MMTV до 10% от транскрипции с дексаметазоном (таблица 1). Этот пониженный уровень транскрипции с RU486 был связан с более быстрыми FRAP, чем с дексаметазоном (фиг. 9c).
Мы также обнаружили, что для промотора MMTV естественный гормон кортикостерон был менее эффективным агонистом, чем синтетический гормон дексаметазон (таблица 1). Снижение уровней транскрипции с кортикостероном снова было связано с более быстрым обменом GFP-GR в матрице, чем с дексаметазоном (рис.9г).
Таким образом, для чисто эндогенного различия (конденсированные и деконденсированные массивы) или для различий, вызванных агонистами или антагонистами, снижение транскрипционной активности сопровождалось более высокой скоростью обмена GFP-GR.
Затем мы исследовали уровни транскрипции после нарушения функции шаперона. После того, как активность hsp90 блокировалась гелданамицином или радициколом, уровни транскрипции падали на матрице MMTV (таблица 1). Это согласуется с предыдущими исследованиями (2), но в этих исследованиях не было ясно, является ли снижение транскрипции результатом неспособности GR связываться с его сайтами-мишенями или даже проникать в ядро.В наших условиях с гелданамицином GR остается связанным с массивом MMTV в течение 10 минут в кортикостероне и в течение 1 часа в дексаметазоне. Однако в эти периоды времени транскрипция все еще значительно снижается, демонстрируя более тонкий эффект на транскрипцию. Одна из возможностей — более высокий обменный курс, измеренный во всех этих случаях.
Наконец, мы измерили уровни транскрипции после ингибирования протеасом. Здесь мы обнаружили зависимость от лиганда. С дексаметазоном уровни транскрипции повысились, но с кортикостероном они снизились (таблица 1).Deroo et al. (8) обнаружили, что длительное лечение (от 22 до 28 часов) MG-132 увеличивало как уровни GR, так и транскрипцию в присутствии дексаметазона. Однако обнаруженные нами изменения транскрипции не были связаны с изменениями уровней белка GFP-GR, поскольку средняя интенсивность флуоресценции GFP-GR в ядрах, обработанных MG-132 в течение 1 часа (97% ± 11%), существенно не различалась. от интенсивности контрольных ядер (100% ± 8%). Несмотря на различия в транскрипции, FRAPs с дексаметазоном или кортикостероном показали замедление и неподвижную фракцию, что указывает на то, что лиганд может иметь совершенно разные эффекты на транскрипцию, когда часть рецепторов становится иммобилизованной.
В итоге, до сих пор мы видели, что во всех случаях изменения обменного курса связаны с изменениями на уровне транскрипции, что позволяет предположить, что они связаны.
ОБСУЖДЕНИЕ
Мы исследовали быстрый обмен GFP-GR в тандемном массиве промоторных сайтов MMTV и предоставили первое понимание его механизма и функции. Наши результаты показывают, что обмен контролируется, по крайней мере частично, балансом между функциями протеасомы и шаперона. Эти две системы часто действуют согласованно, например, обеспечивая баланс между повторной укладкой и деградацией клеточных белков (45).Наши данные предполагают, что сходное уравновешивающее действие осуществляется на промоторе MMTV, при этом шапероны помогают стабилизировать связывание GR, а протеасомы помогают катализировать удаление GR (рис. 10). Как именно достигается этот баланс, неизвестно, но могут быть задействованы такие факторы, как CHIP и BAG-1, которые, как известно, соединяют шаперонную и протеасомную системы (5, 21).
РИС. 10.Модель взаимодействия протеасома-шаперон в матрице MMTV и ассоциации с транскрипцией. Связывание лигандированных GR с MMTV в конечном итоге приводит к образованию комплекса инициации.Формированию комплекса может способствовать более длительное пребывание GR в шаблоне MMTV. Продолжительность занятия GR в шаблоне частично определяется конкуренцией между функциями протеасомы и шаперона. Ингибирование протеасомы способствует занятости GR, что приводит к более медленному FRAP с неподвижной фракцией. Ингибирование шаперона гелданамицином способствует потере GR, что приводит к более быстрой FRAP. Равновесие между этими двумя компонентами помогает установить уровень транскрипции и может частично опосредоваться одним или несколькими факторами, которые, как известно, связывают шаперонную и протеасомную активности (5, 21).
Мы обнаружили, что и шапероны, и протеасомы присутствовали в массиве MMTV и вокруг него, демонстрируя, что оба класса молекул были специфически задействованы в этом сайте. Эти наблюдения согласуются с исследованиями ChIP, демонстрирующими, что либо шапероны (12), либо протеасомы (13, 31) обнаруживаются на сайтах промотора, хотя до сих пор не на одном и том же промоторе.
Функция протеасомы. После нарушения активности протеасомы мы обнаружили, что GFP-GR медленнее восстанавливается на сайтах MMTV и достигает только 95% от своей начальной интенсивности.Остаточные 5% соответствуют неподвижной фракции GFP-GR, которая остается стабильно связанной с промотором после протеасомного ингибирования. Мы пришли к выводу, что удаление по крайней мере части молекул GR обычно требует протеасомы. Поскольку и GR, и различные коактиваторы убиквитинируются (5, 46), потеря GR на промоторе может отражать прямое влияние протеасомы на GR или косвенное последствие потери тесно связанного коактиватора. Наши данные также не различают, включает ли потеря GR обычно деградацию этой 5% фракции GR на промоторе или просто выброс фракции из промотора.Мы не можем исключить возможность того, что обработка протеасом каким-либо иным образом косвенно изменяет подвижность GR, вызывая, например, стрессовую реакцию (40). Однако, используя более короткие процедуры лечения MG-132, которые нарушают преимущественно поведение FRAP в массиве MMTV, мы уменьшаем вероятность того, что наблюдаемые нами эффекты являются неспецифическими. Эта проблема может быть решена более непосредственно в будущих исследованиях путем идентификации и последующего нарушения специфических молекул, которые сочетают активность протеасом с GR.
Известно также, что ингибирование протеасом влияет на ядерную подвижность стероидных рецепторов, индуцируя связывание с ядерным матриксом (8, 34, 43).Однако при более коротких инкубациях с ингибитором протеасомы, который мы использовали, массив MMTV оставался видимым и транскрипция продолжалась, что указывает на то, что GR все еще был связан с ДНК на сайтах MMTV. Наше наблюдение неподвижной фракции в определенных местах после ингибирования дексаметазоном и протеасомами контрастирует с эффектом, обнаруженным для GR в другом месте ядра, где дексаметазон или другие лиганды с высоким сродством ослабляли неподвижную фракцию, вызванную ингибированием протеасом (34). Эти различия показывают, что одна и та же обработка может давать разные эффекты в зависимости от того, является ли сайт связывания специфическим промотором или ядерным матриксом.
Ряд биохимических подходов предполагает роль протеасом в удалении транскрипционного фактора с промоторных сайтов (27). Высокоактивные регуляторы транскрипции, такие как VP16 или myc, должны быть убиквитинированы для индукции транскрипции и после дальнейшего убиквитинирования разрушаются протеасомой (33). Эта деградация может происходить на промоторе, так как она усиливается связыванием ДНК (24). Недавние исследования показали, что медленный цикл ER сопровождается и зависит от цикла протеасомы (31).
Хотя есть интригующее сходство между этими результатами ChIP для ER и нашими данными FRAP для GR, они могут отражать совершенно разные процессы. Это связано с тем, что шкалы времени, исследуемые этими двумя методами, радикально различаются. FRAP GFP-GR на массиве MMTV завершается в течение 1 мин. Таким образом, среднее время пребывания GFP-GR на промоутере может быть не более 1 минуты и вполне может быть намного короче. Такой быстрый обмен не может быть обнаружен с помощью ChIP, поскольку этап фиксации формальдегида в процедуре обычно длится намного дольше (~ 10 мин), чем время цикла GR.Как следствие, ChIP обнаруживает периодичности во временных масштабах на 1-2 порядка больше, чем те, которые обнаруживаются FRAP. Возможно, что гораздо более медленная цикличность GR накладывается на более быструю циклическую смену, которую мы измеряем с помощью FRAP. Доля связанного GR на сайтах MMTV определяется аффинностью связывания GR с этими сайтами. Если это сродство медленно увеличивается, а затем медленно уменьшается, то количество связанного GR будет медленно меняться, даже если отдельные молекулы GR в любой момент будут демонстрировать быстрый обмен, который мы измеряем с помощью FRAP.В принципе, это можно обнаружить в нашей системе живых клеток двумя способами, ни один из которых не был тщательно исследован. Если аффинность связывания GR с MMTV постепенно увеличивается, тогда интенсивность массива должна постепенно увеличиваться, и в то же время скорость FRAP должна постепенно снижаться, поскольку более медленное восстановление отражает связывание с более высокой аффинностью. В общем, одна и та же система может работать как с быстрым, так и с медленным циклом. Быстрые циклы лучше всего обнаруживаются FRAP в целевом массиве промоторов, а медленные циклы лучше наблюдаются с помощью ChIP.
Функция шаперона. Нарушая активность шаперона гелданамицином или радициколом, мы наблюдали ускоренный обмен GFP-GR на сайтах промотора MMTV. Более быстрый обмен GFP-GR был обнаружен мгновенно в присутствии гелданамицина и кортикостерона, что убедительно подтверждает специфический эффект связывания GFP-GR с MMTV.
Нацеливание одного члена шаперонного комплекса, p23, на ряд сайтов промотора GR приводило к потере там связывания GR (12). Основываясь на этом и других наблюдениях, Фриман и Ямамото предположили, что p23 может многократно удалять GR из матрицы и, следовательно, вызывать быстрый процесс обмена GR, наблюдаемый в живых клетках.Однако, если бы это было так, то в простейшем сценарии мы должны были бы наблюдать более медленную FRAP после лечения гелданамицином, поскольку препарат предотвращает связывание p23 с шапероновым комплексом (38). Это, в свою очередь, должно было препятствовать тому, чтобы p23 выбрасывал GR из промотора. Вместо этого мы обнаружили более быстрый обмен GFP-GR, подразумевая, что шапероны обычно стабилизируют связывание GR, а не катализируют его удаление.
Возможен ряд объяснений этого несоответствия. Ключевое различие между двумя исследованиями заключается в том, что мы нарушили функцию hsp90, тогда как Фриман и Ямамото изменили активность p23.Хотя p23 и hsp90 обычно действуют согласованно внутри комплекса шаперонов, они не могут действовать вместе в промоторе. Если это так, то лечение гелданамицином будет напрямую влиять на hsp90, но не на p23. Затем наши данные FRAP в присутствии гелданамицина означают, что hsp90 необходим для стабилизации связывания GR на промоторе, тогда как данные ChIP Фримена и Ямамото предполагают, что p23 необходим для его дестабилизации. Такие антагонистические действия между hsp90 и p23 могут играть роль в установлении времен резидентности GR.В более общем плане должен существовать тонкий баланс между функциями шаперона и протеасомы в промоторе MMTV. Таким образом, даже если лечение гелданамицином также нарушает функцию p23 в MMTV, возможно, что конкретный метод, используемый для изменения активности p23, может изменить равновесие между шаперонами и протеасомами в ту или иную сторону, что приведет к противоположным эффектам на связывание GR. Эти несоответствия, вероятно, будут устранены путем выяснения молекулярного механизма, который устанавливает взаимодействие между шаперонами и протеасомами на промоторе MMTV.В настоящее время мы можем с уверенностью сказать, что как наши результаты, так и результаты Фримена и Ямамото строго подтверждают роль шаперонов в связывании GR на промоторе.
Функция быстрого обмена. Было высказано предположение, что цикл фактора транскрипции на промоторе является механизмом определения изменений окружающей среды, например, концентрации лиганда (12, 22, 31, 37). Через 5 минут после удаления кортикостерона мы обнаружили, что транскрипция с промотора MMTV практически прекратилась. Это согласуется с моделью восприятия для обмена, поскольку время цикла GR ~ 1 мин достаточно быстро, чтобы показать ответ в течение 5 минут после падения концентрации гормона.Обратите внимание, однако, что такое быстрое изменение уровней гормонов может быть обнаружено только с помощью механизма быстрого обмена, а не по более длительному времени цикла, наблюдаемому для ER с помощью ChIP (31, 37).
Помимо возможной роли в обнаружении измененных уровней гормонов, мы обнаружили связь между уровнем транскрипции и скоростью обмена. Во всех изученных до сих пор случаях, когда менялись уровни транскрипции, менялся и обменный курс. В настоящее время наши данные подтверждают простую форму связывания, а именно, что более медленный обмен связан с большей транскрипцией.Это могло произойти, потому что более длительное время пребывания GR может увеличить шансы на успешную загрузку полимеразы. Мы наблюдали эту корреляцию между скоростью обмена и уровнем транскрипции в семи из восьми двусторонних сравнений (таблица 1).
Единственным исключением были ингибитор протеасом MG-132 и лиганд кортикостерон. Эта комбинация приводила к более медленному обмену и снижению транскрипции, тогда как MG-132 и дексаметазон приводили к более медленному обмену и повышенной транскрипции.Одним из объяснений этого несоответствия является врожденная разница между временем пребывания лиганда, связывающегося с GR с дексаметазоном и кортикостероном. После ингибирования протеасомы MG-132 часть GR остается на матрице.

Об авторе